Гаплогруппа R1b
| Гаплогруппа R1b | |
|---|---|
.png/1280px-Haplogrupo_R1b_(ADN-Y).png) | |
| Возможное время происхождения | Вероятно, вскоре после R1, возможно, между 18 000 и 14 000 годами до н.э. [1] |
| Возможное место происхождения | Западная Азия , Северная Евразия или Восточная Европа [2] |
| Предок | Р1 |
| Потомки |
|
| Определение мутаций | М343 |
.png){kind=link}
Гаплогруппа R1b ( R-M343 ), ранее известная как Hg1 и Eu18 , является гаплогруппой Y-хромосомы человека .
Это наиболее часто встречающаяся отцовская линия в Западной Европе , а также в некоторых частях России (например, у башкир ) и по всему Сахелю в Центральной Африке , а именно: в Камеруне , Чаде , Гвинее , Мавритании , Мали , Нигере , Нигерии и Сенегале (сосредоточена в некоторых частях Чада с концентрацией в племени хауса и среди говорящих на чадском языке этнических групп Камеруна).
Клада также присутствует с меньшей частотой в Восточной Европе , Западной Азии , Центральной Азии, а также в некоторых частях Северной Африки , Южной Азии и Центральной Азии .
R1b имеет две основные ветви: R1b1-L754 и R1b2-PH155. R1b1-L754 имеет два основных субклада: R1b1a1b-M269, который преобладает в Западной Европе, и R1b1b-V88, который сегодня распространен в некоторых частях Центральной Африки. Другая ветвь, R1b2-PH155, настолько редка и широко распространена, что трудно сделать какие-либо выводы о ее происхождении. Она была обнаружена в Бахрейне , Индии , Непале , Бутане , Ладакхе , Таджикистане , Турции и Западном Китае .
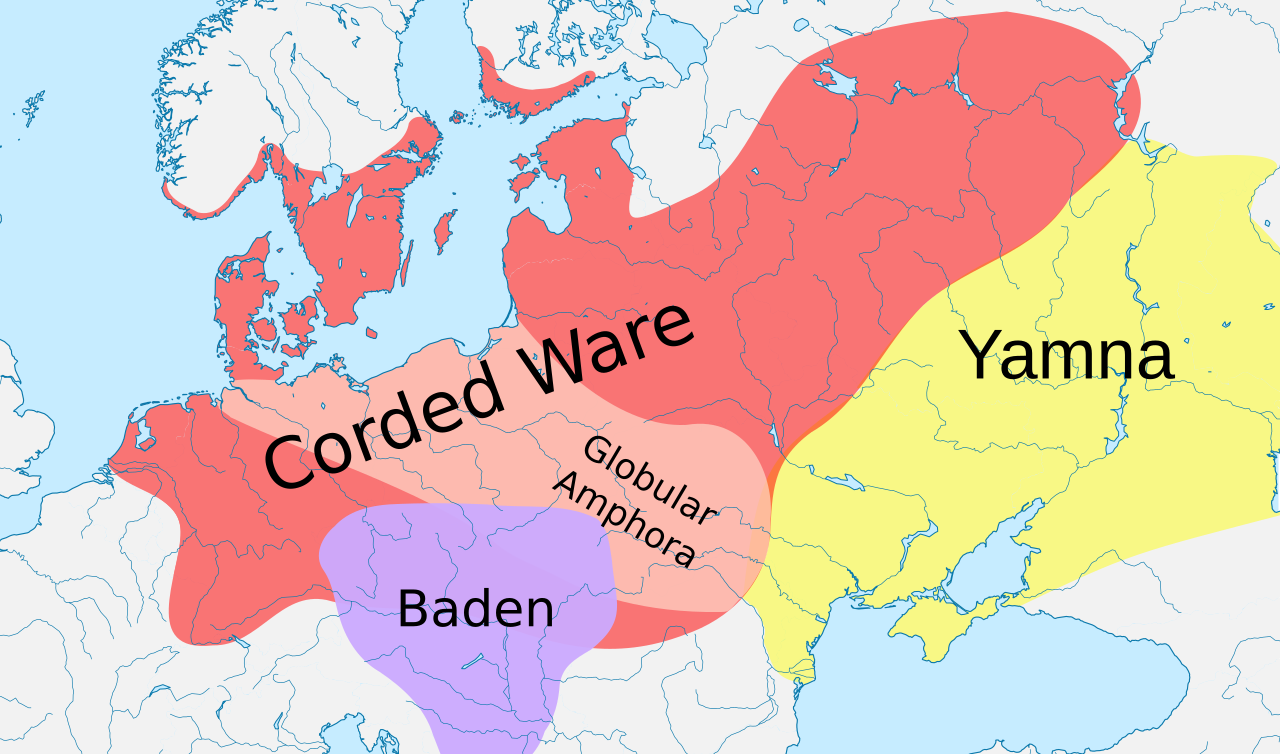
Согласно исследованиям древней ДНК, большинство линий R1a и R1b распространились из Понтийской степи вместе с индоевропейскими языками . [2] [3] [4] [5] [6]
Происхождение и распространение
{kind=link}
Возраст R1 был оценен Татьяной Карафет и соавторами (2008) как 12 500–25 700 лет назад , и, скорее всего, он появился около 18 500 лет назад. [7] Поскольку самый ранний известный пример датируется примерно 14 000 лет назад и относится к R1b1 (R-L754), [1] R1b должен был возникнуть относительно скоро после появления R1.
Ранние останки человека, в которых обнаружены носители R1b, включают:
- Виллабруна 1 (особь I9030), западный охотник-собиратель (WHG), найденный в эпиграветтской культуре в долине Цизмон (современный Венето , Италия), живший около 14000 лет до нашей эры и принадлежавший к R1b1a. [1] [8]
- Несколько мужчин мезолита Железных Ворот на Балканах , похороненных между 11200 и 8200 годами до нашей эры, несли R1b1a1a. Было установлено, что эти люди в основном имели происхождение от WHG, с небольшой примесью восточных охотников-собирателей (EHG). [9]
- Несколько мужчин мезолитической культуры Кунда и неолитической культуры Нарва , похороненных в могильнике Звейниеки на территории современной Латвии около 9500–6000 лет до н. э., имели R1b1b. [10] [11] Было установлено, что эти люди в основном имели происхождение от WHG с небольшой примесью EHG. [10]
- Несколько мужчин эпохи мезолита и неолита, захороненных в Дериивке и Васильевке на территории современной Украины около 9500-7000 лет до нашей эры, имели аллель R1b1a. [12] Эти люди в основном имели происхождение от EHG со значительной примесью WHG. [13]
- Мужчина WHG, похороненный в Островул Корбули, Румыния, около 8700 г. до н.э., имел R1b1c. [14] [15] [16]
- Мужчина, похороненный в Лепенски Вир , Сербия, около 8200-7900 лет до нашей эры, имел генетический материал R1b1a. [17]
- EHG, захороненный около Самары , Россия, 7500 лет до нашей эры, нес R1b1a1a. [18]
- Энеолитический мужчина , похороненный в Хвалынске , Россия, около 7200-6000 лет до нашей эры, носил R1b1a. [19]
- Неолитический мужчина , захороненный в Эльс-Трокс , Испания, около 7178-7066 гг. до н. э., который, возможно, принадлежал к эпикардиальной культуре [20] , оказался носителем R1b1. [21] [22] [23]
- Поздний халколитический мужчина, похороненный в Смядово , Болгария, около 6500 г. до н. э., носил R1b1a. [24]
- Мужчина раннего медного века, захороненный в Каннас-ди-Сотто, Карбония, Сардиния, около 6450 г. до н.э., имел генетический материал R1b1b2. [25]
- Мужчина из группы Баальберге в Центральной Европе, похороненный около 5600 г. до н.э., имел аллель R1b1a. [26]
- Мужчина ботайской культуры в Центральной Азии, похороненный около 5500 лет до нашей эры, имел R1b1a1 (R1b-M478). [27]
- Было обнаружено, что 7 мужчин, прошедших тестирование в ямной культуре , принадлежат к субкладу M269 гаплогруппы R1b. [28]
R1b является субкладом в пределах «макрогаплогруппы » K ( M9), наиболее распространенной группы человеческих мужских линий за пределами Африки. Считается, что K возникла в Азии (как и в случае с еще более ранней предковой гаплогруппой F (F-M89). Карафет Т. и др. (2014) предположили, что «быстрый процесс диверсификации K-M526 , вероятно, произошел в Юго-Восточной Азии , с последующей экспансией на запад предков гаплогрупп R и Q ». [29] Однако самый старый пример R* был обнаружен в древнем североевразийском образце из Сибири ( мальтинский мальчик , 24 000 лет назад), а его предшественник P1 был обнаружен в другом древнем североевразийском образце из северной Сибири ( Яна RHS ), датируемом примерно 31 600 лет назад. [30]
Три генетических исследования в 2015 году подтвердили гипотезу Кургана Марии Гимбутас относительно прародины праиндоевропейцев . Согласно этим исследованиям, гаплогруппы R1b-M269 и R1a, в настоящее время наиболее распространенные в Европе (R1a также распространена в Южной Азии), могли распространиться из западной евразийской степи вместе с индоевропейскими языками ; они также обнаружили аутосомный компонент, присутствующий у современных европейцев, который отсутствовал у неолитических европейцев , который мог быть введен с отцовскими линиями R1b и R1a, а также с индоевропейскими языками. [2] [3] [4]
{kind=link}

Анализ древней Y-ДНК из останков ранних неолитических поселений культуры линейной керамики в Центральной и Северной Европе пока не выявил мужчин, принадлежащих к гаплогруппе R1b-M269. [31] [32] Олальде и др. (2017) прослеживают распространение гаплогруппы R1b-M269 в Западной Европе, в частности в Британии, до распространения культуры кубков , с внезапным появлением множества гаплогрупп R1b-M269 в Западной Европе примерно 5000–4500 лет назад в течение раннего бронзового века. [33]
Структура
Внешняя филогения R1b
Более широкая гаплогруппа R (M207) является первичным субкладом гаплогруппы P1 (M45), которая сама является первичной ветвью P (P295), также известной как гаплогруппа K2b2. Таким образом, R-M207 является вторичной ветвью K2b (P331) и прямым потомком K2 (M526).
- Филогенетика в пределах K2b
- P P295/PF5866/S8 (также известный как K2b2 ).
Внутренняя структура R1b
Такие названия, как R1b, R1b1 и т. д., являются филогенетическими (т. е. «генеалогическими») названиями, которые четко указывают на их место в разветвлении гаплогрупп или филогенетическом дереве. Альтернативный способ именования тех же гаплогрупп и субкладов относится к их определяющим мутациям SNP : например, R-M343 эквивалентен R1b. [34] Филогенетические названия меняются с новыми открытиями , и названия на основе SNP впоследствии реклассифицируются в филогенетическом дереве. В некоторых случаях SNP оказывается ненадежным в качестве определяющей мутации, и название на основе SNP полностью удаляется. Например, до 2005 года R1b был синонимом R-P25, который позже был реклассифицирован как R1b1; в 2016 году R-P25 был полностью удален в качестве определяющего SNP из-за значительной скорости обратной мутации. [35] (Ниже приведена основная схема R1b согласно дереву ISOGG по состоянию на 30 января 2017 года. [36] )
|
Географическое распределение
R1b* (R-M343*)
В рецензируемой литературе не сообщалось о подтвержденных случаях R1b* (R-M343*), то есть R1b (xR1b1, R1b2), также известного как R-M343 (xL754, PH155) .
- Р-М343 (xM73, M269, V88)
В ранних исследованиях, поскольку R-M269, R-M73 и R-V88 являются наиболее распространенными формами R1b, примеры R1b (xM73, xM269) иногда предполагались как обозначающие базальные примеры «R1b*». [35] Однако, хотя парагруппа R-M343 (xM73, M269, V88) встречается редко, это не исключает членства в редких и/или впоследствии открытых, относительно базальных субкладах R1b, таких как R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2) или R-PH155 (R1b2).
Популяция, которая, как полагают, имеет самую высокую долю R-M343 (xM73, M269, V88), - это курды юго-восточного Казахстана - 13%. [41] Однако совсем недавно крупное исследование вариаций Y-хромосомы в Иране выявило R-M343 (xV88, M73, M269) на уровне 4,3% среди иранских субпопуляций. [42]
Остается вероятность, что некоторые или даже большинство из этих случаев могут быть R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2), R-PH155 (R1b2), R1b* (R-M343*), R1a* (R-M420*), иным образом недокументированной ветвью R1 (R-M173) и/или обратными мутациями маркера от положительного к отрицательному предковому состоянию [43] и, следовательно, представлять собой недокументированные субклады R1b.
Компиляцию предыдущих исследований, касающихся распределения R1b, можно найти в работе Cruciani et al. (2010). [44] Она обобщена в следующей таблице. (Cruciani не включил некоторые исследования, предполагающие даже более высокие частоты R1b1a1b [R-M269] в некоторых частях Западной Европы.)
| Континент | Область | Размер выборки | Всего R1b | R-P25 (ненадежный маркер для R1b1*) | Р-В88 (R1b1b) | Р-М269 (R1b1a1a2) | Р-М73 (R1b1a1a1) |
|---|---|---|---|---|---|---|---|
| Африка | Северная Африка | 691 | 5.9% | 0.0% | 5.2% | 0,7% | 0.0% |
| Африка | Центральный Сахельский регион | 461 | 23.0% | 0.0% | 23.0% | 0.0% | 0.0% |
| Африка | Западная Африка | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Африка | Восточная Африка | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Африка | Южная Африка | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Европа | Западные европейцы | 465 | 57,8% | 0.0% | 0.0% | 57,8% | 0.0% |
| Европа | Северо-западные европейцы | 43 | 55,8% | 0.0% | 0.0% | 55,8% | 0.0% |
| Европа | Центральная Европа | 77 | 42,9% | 0.0% | 0.0% | 42,9% | 0.0% |
| Европа | Северо-восточные европейцы | 74 | 1,4% | 0.0% | 0.0% | 1,4% | 0.0% |
| Европа | русские | 60 | 6.7% | 0.0% | 0.0% | 6.7% | 0.0% |
| Европа | Восточноевропейцы | 149 | 20,8% | 0.0% | 0.0% | 20,8% | 0.0% |
| Европа | Юго-восточные европейцы | 510 | 13,1% | 0.0% | 0,2% | 12,9% | 0.0% |
| Азия | Западные азиаты | 328 | 5.8% | 0.0% | 0,3% | 5,5% | 0.0% |
| Азия | Южные азиаты | 288 | 4,8% | 0.0% | 0.0% | 1,7% | 3.1% |
| Азия | Юго-восточные азиаты | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Азия | Северо-восточные азиаты | 30 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Азия | Восточные азиаты | 156 | 0,6% | 0.0% | 0.0% | 0,6% | 0.0% |
| Общий | 5326 | ||||||
R1b (R-L278)
R-L278 среди современных мужчин попадает в субклады R-L754 и R-PH155, хотя возможно, что могут существовать некоторые очень редкие R-L278*, поскольку не все примеры были протестированы для обеих ветвей. Примеры также могут существовать в древней ДНК, хотя из-за низкого качества часто невозможно сказать, несли ли древние мутации, которые определяют субклады.
Некоторые примеры, описанные в более старых статьях, например, два, найденные в Турции, [34] теперь считаются в основном относящимися к недавно обнаруженному подкладу R1b1b (R-V88). Поэтому большинство примеров R1b попадают в подклады R1b1b (R-V88) или R1b1a (R-P297). Cruciani et al. в большом исследовании 2010 года обнаружили 3 случая среди 1173 итальянцев, 1 из 328 жителей Западной Азии и 1 из 156 жителей Восточной Азии. [44] Varzari обнаружил 3 случая в Украине , в исследовании 322 человек из региона Днестровско - Карпатских гор , которые были положительными по P25, но отрицательными по M269. [45] Случаи из более старых исследований в основном из Африки, Ближнего Востока или Средиземноморья и обсуждаются ниже как вероятные случаи R1b1b (R-V88).
R1b1 (R-L754)
R-L754 содержит подавляющее большинство R1b. Единственный известный пример R-L754* (xL389, V88) также является самым ранним известным носителем R1b: « Виллабруна 1 », живший около 14 000 лет назад (северо-восток Италии). Виллабруна 1 принадлежал к эпиграветтской культуре.
R1b1a (R-L389)
R-L389, также известный как R1b1a (L388/PF6468, L389/PF6531), содержит очень распространенный субклад R-P297 и редкий субклад R-V1636. Неизвестно, относятся ли все ранее зарегистрированные R-L389* (xP297) к R-V1636 или нет.
R1b1a1 (R-P297)
Маркер SNP P297 был признан в 2008 году предковым для значимых субкладов M73 и M269, объединив их в один кластер. [7] Ему было дано филогенетическое название R1b1a1a (а ранее R1b1a).
Большинство евразийских R1b попадает в этот субклад, представляя очень большую современную популяцию. Хотя сам P297 еще не был тщательно протестирован, эта же популяция была относительно хорошо изучена с точки зрения других маркеров. Поэтому ветвление внутри этого клада может быть объяснено ниже с относительно высокой степенью детализации.
R1b1a1a (R-M73)
Малярчук и др. (2011) обнаружили R-M73 у 13,2% (5/38) шорцев, 11,4% (5/44) телеутов, 3,3% (2/60) калмыков, 3,1% (2/64) хакасов, 1,9% (2/108) тувинцев и 1,1% (1/89) алтайцев. [46] Калмыки, тувинцы и алтайцы принадлежат к кластеру Y-STR, обозначенному DYS390=19, DYS389=14-16 (или 14–15 в случае алтайца) и DYS385=13-13.
Дулик и др. (2012) обнаружили R-M73 у 35,3% (6/17) выборки кумандинцев Республики Алтай в России. [47] Трое из этих шести кумандинцев имеют идентичный 15-локусный гаплотип Y-STR, а еще двое отличаются только по локусу DYS458, имея DYS458=18 вместо DYS458=17. Эта пара кумандинских гаплотипов R-M73 напоминает гаплотипы двух калмыков, двух тувинцев и одного алтайца, чья Y-ДНК была проанализирована Малярчуком и др. (2011). Оставшийся кумандин R-M73 имеет гаплотип Y-STR, который резко отличается от гаплотипов других кумандинцев R-M73, напоминая вместо этого гаплотипы пяти шорцев, пяти телеутов и двух хакасов. [46]
Хотя ранние исследования R-M73 утверждали, что он был значительно представлен среди хазарейцев Афганистана и башкир Уральских гор, это, по-видимому, было опровергнуто. Например, подтверждающий материал из исследования 2010 года Бехара и др. предположил, что Сенгупта и др. (2006) могли неправильно идентифицировать хазарейцев, которые вместо этого принадлежали к «PQR2», а не к «R(xR1a)». [48] [41] [49] Однако отнесение Y-ДНК этих хазарейцев к категории «PQR2» Бехаром и др. (2010), вероятно, можно объяснить привычкой, которая была популярна некоторое время, маркировать R-M269 как «R1b» или «R(xR1a)», при этом любые члены R-M343 (xM269) помещались в полифилетическую, всеобъемлющую категорию «R*» или «P». Майрес и др. (2011), Ди Кристофаро и др. (2013) и Липпольд и др. (2014) согласны с тем, что Y-ДНК 32% (8/25) выборки HGDP пакистанских хазарейцев должна принадлежать к гаплогруппе R-M478/M73. [41] [50] [51] Аналогичным образом, было обнаружено, что большинство башкирских мужчин принадлежат к U-152 (R1b1a1a2a1a2b), а некоторые, в основном из юго-восточного Башкортостана, принадлежали к гаплогруппе Q-M25 (Q1a1b), а не к R1b; напротив, Майрес и др. (2011) обнаружили высокую частоту R-M73 среди своей выборки башкир из юго-восточного Башкортостана (77/329 = 23,4% R1b-M73), что согласуется с более ранним исследованием башкир. [41] Помимо высокой частоты R-M73 у юго-восточных башкир, Майрес и др. также сообщалось об обнаружении R-M73 в следующих образцах: 10,3% (14/136) балкарцев с северо-западного Кавказа, 9,4% (8/85) образцов HGDP с северного Пакистана (это вышеупомянутые пакистанские хазарейцы), 5,8% (4/69) карачаевцев с северо-западного Кавказа, 2,6% (1/39) татар из Башкортостана, 1,9% (1/54) башкир с юго-западного Башкортостана, 1,5% (1/67) мегрелов с южного Кавказа, 1,4% (1/70) башкир с северного Башкортостана, 1,3% (1/80) татар из Казани, 1,1% (1/89) образца из Каппадокии, Турция, 0,7% (1/141) кабардинцев с северо-запада Кавказ, 0,6% (3/522) выборки из Турции и 0,38% (1/263) русских из Центральной России. [41]
Помимо вышеупомянутых пакистанских хазарейцев, Ди Кристофаро и др. (2013) обнаружили R-M478/M73 у 11,1% (2/18) монголов из центральной Монголии, 5,0% (1/20) киргизов с юго-запада Кыргызстана, 4,3% (1/23) монголов с юго-востока Монголии, 4,3% (4/94) узбеков из Джаузджана, Афганистан, 3,7% (1/27) иранцев из Гиляна , 2,5% (1/40) киргизов из центрального Кыргызстана, 2,1% (2/97) монголов с северо-запада Монголии и 1,4% (1/74) туркмен из Джаузджана, Афганистан. [50] Монголы, а также человек с юго-запада Кыргызстана, человек из Гиляна и один из узбеков из Джаузджана принадлежат к тому же кластеру гаплотипа Y-STR, что и пять из шести членов кумандинской расы R-M73, изученных Дуликом и др. (2012). Наиболее отличительным значением Y-STR этого кластера является DYS390=19. [41]
Карафет и др. (2018) обнаружили R-M73 у 37,5% (15/40) выборки телеутов из Беково Кемеровской области, 4,5% (3/66) выборки уйгуров из Синьцзян-Уйгурского автономного района, 3,4% (1/29) выборки казахов из Казахстана, 2,3% (3/129) выборки селькупов, 2,3% (1/44) выборки туркмен из Туркменистана и 0,7% (1/136) выборки иранцев из Ирана. [52] Четверо из этих людей (один из телеутов, один из уйгуров, казах и иранец), по-видимому, принадлежат к вышеупомянутому кластеру, обозначенному DYS390=19 (кумандино-монгольский кластер R-M73); Телеуты и уйгуры также разделяют модальные значения в локусах DYS385 и DYS389. Иранский язык отличается от модального для этого кластера тем, что имеет 13-16 (или 13–29) в DYS389 вместо 14-16 (или 14–30). Казахский язык отличается от модального языка тем, что имеет 13–14 в DYS385 вместо 13-13. Остальные четырнадцать телеутов и три селькупа, по-видимому, принадлежат к телеутско-шорско-хакасскому кластеру R-M73 из набора данных Малярчука и др. (2011); этот кластер имеет модальные значения DYS390=22 (но 21 в случае двух телеутов и одного хакаса), DYS385=13-16 и DYS389=13-17 (или 13–30, но 14–31 в случае одного селькупа).
Казахстанская статья, опубликованная в 2017 году, обнаружила гаплогруппу R1b-M478 Y-ДНК у 3,17% (41/1294) выборки казахов из Казахстана, причем эта гаплогруппа наблюдалась с частотой выше средней среди членов племен кыпшак (12/29 = 41,4%), ысты (6/57 = 10,5%), конгырат (8/95 = 8,4%), ошакты (2/29 = 6,9%), керей (1/28 = 3,6%) и джетыру (3/86 = 3,5%) . [ 53] Китайская статья, опубликованная в 2018 году, обнаружила гаплогруппу R1b-M478 Y-ДНК у 9,2% (7/76) выборки доланских уйгуров из поселка Хорикол, уезд Ават , Синьцзян. [54]
R1b1a1b (R-M269)
R-M269, или R1b1a1b (по состоянию на 2018 год) среди других названий, [55] в настоящее время является наиболее распространенной линией Y-ДНК у европейских мужчин. Ее носителями являются, по оценкам, 110 миллионов мужчин в Европе. [56]
{kind=link}

R-M269 получил значительный научный и общественный интерес из-за его возможной связи с индоевропейской экспансией в Европе. В частности, было обнаружено, что субклад R-Z2103 преобладает в древней ДНК, связанной с культурой Ямна . [2] Все семь индивидуумов в одном были определены как принадлежащие к субкладу R1b-M269. [2]
Более ранние исследования, опубликованные до того, как исследователи смогли изучить ДНК древних останков, предполагали, что R-M269, вероятно, возник в Западной Азии и присутствовал в Европе к периоду неолита. [36] [41] [57] [58] Но результаты, основанные на реальной древней ДНК, отметили, что в Европе до бронзового века наблюдался дефицит R-M269, [2] а распределение субкладов в Европе в значительной степени обусловлено различными миграциями бронзового и железного веков . Аналогичным образом, самые старые образцы, классифицированные как принадлежащие к R-M269, были найдены в Восточной Европе и Понтийско-Каспийской степи, а не в Западной Азии. Западноевропейские популяции разделены между субкладами R-P312/S116 и R-U106/S21 R-M412 (R-L51).
Распространение R-M269 в Европе увеличивается по частоте с востока на запад. Он достигает пика на национальном уровне в Уэльсе с показателем 92%, 82% в Ирландии , 70% в Шотландии , 68% в Испании , 60% во Франции (76% в Нормандии ), около 60% в Португалии , [41] 50% в Германии , 50% в Нидерландах , 47% в Италии , [59] 45% в Восточной Англии и 42% в Исландии . R-M269 достигает уровней до 95% в некоторых частях Ирландии. Он также был обнаружен с более низкими частотами по всей центральной Евразии , [60] но с относительно высокой частотой среди башкир Пермского края (84,0%). [61] Этот маркер присутствует в Китае и Индии с частотами менее одного процента. В Северной Африке и прилегающих островах, в то время как R-V88 (R1b1b) представлен более сильно, R-M269, по-видимому, присутствовал с древности. R-M269 был обнаружен, например, с частотой ~44% среди останков, датируемых 11-13 веками в Пунта-Асуль , на Канарских островах . Эти останки были связаны с бимбаче (или бимапе), подгруппой гуанчей. [62] У ныне живущих мужчин он достигает пика в некоторых частях Северной Африки, особенно в Алжире , с частотой 10%. [63] В странах Африки к югу от Сахары пик R-M269, по-видимому, наблюдается в Намибии , с частотой 8% среди мужчин гереро . [64] В Западной Азии R-M269 был зарегистрирован у 40% армянских мужчин и более чем у 35% туркменских мужчин. [65] [66] (В таблице ниже более подробно перечислены частоты M269 в регионах Азии, Европы и Африки.)
Помимо базального R-M269*, который не дивергировал, существуют (по состоянию на 2017 год) две основные ветви R-M269:
- Р-Л23 (R1b1a1b1; L23/PF6534/S141) и
- R-PF7558 (R1b1a1b2; PF7558/PF7562.)
R-L23 (Z2105/Z2103; также известный как R1b1a1b1) был зарегистрирован среди народов Идель -Урала (по Трофимовой и др., 2015): 21 из 58 (36,2%) башкир Бурзянского района , 11 из 52 (21,2%) удмуртов , 4 из 50 (8%) коми , 4 из 59 (6,8%) мордвы , 2 из 53 (3,8%) бесермян и 1 из 43 (2,3%) чувашей были носителями R1b-L23. [67]
Субклады внутри парагруппы R-M269(xL23) – то есть R-M269* и/или R-PF7558 – по-видимому, встречаются с наибольшей частотой в центральных Балканах , особенно в Косово с показателем 7,9%, Северной Македонии с показателем 5,1% и Сербии с показателем 4,4%. [41] В отличие от большинства других регионов со значительным процентом R-L23, Косово , Польша и башкиры юго-востока Башкортостана отличаются высоким процентом R-L23 (xM412) – с показателем 11,4% (Косово), 2,4% (Польша) и 2,4% на юго-востоке Башкортостана. [41] (Эта башкирская популяция также примечательна своим высоким уровнем R-M73 (R1b1a1a1) - 23,4%. [41] ) Пять человек из 110, протестированных в Араратской долине Армении , принадлежали к R-M269(xL23) и 36 к R-L23*, при этом ни один из них не принадлежал к известным субкладам L23. [68]
В 2009 году ДНК, извлеченная из бедренных костей 6 скелетов в раннесредневековом захоронении в Эргольдинге (Бавария, Германия), датируемом примерно 670 годом н. э., дала следующие результаты: было обнаружено, что 4 из них относятся к гаплогруппе R1b с наиболее близкими соответствиями в современных популяциях Германии, Ирландии и США, а 2 относятся к гаплогруппе G2a . [69]
Ниже приводится краткое изложение большинства исследований, в которых конкретно изучался M269, показывающее его распространение (в процентах от общей численности населения) в Европе, Северной Африке , на Ближнем Востоке и в Центральной Азии вплоть до Китая и Непала .
Филогения R-M269 согласно ISOGG 2017:
| М269/ПФ6517 |
| |||||||||||||||||||||||||||||||||||||||||||||
R1b1b (R-V88)
R1b1b (PF6279/V88; ранее R1b1a2) определяется наличием маркера SNP V88, открытие которого было объявлено в 2010 году Кручиани и др. [44] Помимо людей в Южной Европе и Западной Азии, большинство R-V88 было обнаружено в Сахеле, особенно среди популяций, говорящих на афразийских языках чадской ветви.
На основании подробного филогенетического анализа Д'Атанасио и др. (2018) предположили, что R1b-V88 возникла в Европе около 12 000 лет назад и перешла в Северную Африку между 8000 и 7000 лет назад, в период « Зеленой Сахары ». R1b-V1589, основной субклад в пределах R1b-V88, подвергся дальнейшему расширению около 5500 лет назад, вероятно, в районе бассейна озера Чад, откуда некоторые линии пересекли Сахару и попали в Северную Африку. [70]
Маркус и др. (2020) приводят убедительные доказательства в пользу этой предлагаемой модели транссахарского движения с севера на юг: самые ранние базальные гаплогруппы R1b-V88 обнаружены у нескольких восточноевропейских охотников-собирателей около 11 000 лет назад. Затем гаплогруппа, по-видимому, распространилась с экспансией неолитических земледельцев , которые основали сельское хозяйство в Западном Средиземноморье примерно 7500 лет назад. Гаплогруппы R1b-V88 были обнаружены у древних неолитических людей в Германии, центральной Италии, Иберии и, особенно часто, на Сардинии. Часть ветви, ведущей к современным африканским гаплогруппам (V2197), уже была получена у неолитических европейских людей из Испании и Сардинии, что обеспечивает дополнительную поддержку транссахарского движения с севера на юг. [71] [72] [73] Европейское аутосомное происхождение, гаплогруппы мтДНК и аллели устойчивости к лактазе также были выявлены в африканских популяциях, которые несут R1b-V88 с высокой частотой, таких как фулани и тубу . [74] [75] [72] [76] Присутствие европейских неолитических земледельцев в Африке дополнительно подтверждается образцами из Марокко, датируемыми примерно 5400 г. до н.э. [77] [78]
Исследования 2005–08 гг. выявили высокий уровень «R1b*» в Иордании , Египте и Судане . [79] [64] [80] [примечание 1] Последующие исследования Майреса и др. (2011) показывают, что соответствующие образцы, скорее всего, принадлежат к подкладу R-V88. Согласно Майресу и др. (2011), это может быть объяснено обратной миграцией из Азии в Африку людей, несущих R1b. [41] [примечание 2]
В отличие от других исследований, Шрайнер и Ротими (2018) связали появление R1b в Чаде с недавними перемещениями арабов баггара . [81]
| Область | Население | Страна | Язык | Размер выборки | Общий % | R1b1b (R-V88) | R1b1a1b (R-M269) | R1b1b* (R-V88*) | R1b1b2a2a1 (R-V69) |
|---|---|---|---|---|---|---|---|---|---|
| Северная Африка | Композитный | Марокко | АА | 338 | 0.0% | 0,3% | 0,6% | 0,3% | 0.0% |
| Северная Африка | Мозабитские берберы | Алжир | АА/берберский | 67 | 3.0% | 3.0% | 0.0% | 3.0% | 0.0% |
| Северная Африка | Северные египтяне | Египет | АА/семитский | 49 | 6.1% | 4.1% | 2.0% | 4.1% | 0.0% |
| Северная Африка | Берберы из Сивы | Египет | АА/берберский | 93 | 28.0% | 26,9% | 1.1% | 23,7% | 3.2% |
| Северная Африка | Бахария | Египет | АА/семитский | 41 | 7.3% | 4,9% | 2.4% | 0.0% | 4,9% |
| Северная Африка | Оазис Гурна | Египет | АА/семитский | 34 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Северная Африка | Южные египтяне | Египет | АА/семитский | 69 | 5.8% | 5.8% | 0.0% | 2,9% | 2,9% |
| С Африка | Сонгай | Нигер | NS/Сонгай | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| С Африка | Фульбе | Нигер | Северная Каролина/Атлантика | 7 | 14,3% | 14,3% | 0.0% | 14,3% | 0.0% |
| С Африка | Туареги | Нигер | АА/берберский | 22 | 4,5% | 4,5% | 0.0% | 4,5% | 0.0% |
| С Африка | Нгамбаи | Чад | NS/Суданские | 11 | 9.1% | 9.1% | 0.0% | 9.1% | 0.0% |
| С Африка | хауса | Нигерия (Север) | АА/Чадский | 10 | 20.0% | 20.0% | 0.0% | 20.0% | 0.0% |
| С Африка | Фульбе | Нигерия (Север) | Северная Каролина/Атлантика | 32 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| С Африка | йоруба | Нигерия (Юг) | NC/Дефоид | 21 | 4,8% | 4,8% | 0.0% | 4,8% | 0.0% |
| С Африка | Оулдеме | Камерун (северный) | АА/Чадский | 22 | 95,5% | 95,5% | 0.0% | 95,5% | 0.0% |
| С Африка | Мада | Камерун (северный) | АА/Чадский | 17 | 82,4% | 82,4% | 0.0% | 76,5% | 5.9% |
| С Африка | Мафа | Камерун (северный) | АА/Чадский | 8 | 87,5% | 87,5% | 0.0% | 25.0% | 62,5% |
| С Африка | Гуизига | Камерун (северный) | АА/Чадский | 9 | 77,8% | 77,8% | 0.0% | 22.2% | 55,6% |
| С Африка | Даба | Камерун (северный) | АА/Чадский | 19 | 42.1% | 42.1% | 0.0% | 36,8% | 5.3% |
| С Африка | Гуидар | Камерун (северный) | АА/Чадский | 9 | 66,7% | 66,7% | 0.0% | 22.2% | 44,4% |
| С Африка | Масса | Камерун (северный) | АА/Чадский | 7 | 28,6% | 28,6% | 0.0% | 14,3% | 14,3% |
| С Африка | Другие чадские | Камерун (северный) | АА/Чадский | 4 | 75.0% | 75.0% | 0.0% | 25.0% | 50.0% |
| С Африка | Арабы Шува | Камерун (северный) | АА/семитский | 5 | 40.0% | 40.0% | 0.0% | 40.0% | 0.0% |
| С Африка | Канури | Камерун (северный) | NS/Сахарский | 7 | 14,3% | 14,3% | 0.0% | 14,3% | 0.0% |
| С Африка | Фульбе | Камерун (северный) | Северная Каролина/Атлантика | 18 | 11.1% | 11.1% | 0.0% | 5.6% | 5.6% |
| С Африка | Мунданг | Камерун (северный) | Северная Каролина/Адамава | 21 | 66,7% | 66,7% | 0.0% | 14,3% | 52.4% |
| С Африка | Фали | Камерун (северный) | Северная Каролина/Адамава | 48 | 20,8% | 20,8% | 0.0% | 10,4% | 10,4% |
| С Африка | Тали | Камерун (северный) | Северная Каролина/Адамава | 22 | 9.1% | 9.1% | 0.0% | 4,5% | 4,5% |
| С Африка | Мбум | Камерун (северный) | Северная Каролина/Адамава | 9 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| С Африка | Композитный | Камерун (Южный) | Северная Каролина/Банту | 90 | 0.0% | 1.1% | 0.0% | 1.1% | 0.0% |
| С Африка | Пигмеи Биака | МАШИНА | Северная Каролина/Банту | 33 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Западная Африка | Композитный | — | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Восточная Африка | Композитный | — | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Южная Африка | Композитный | — | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Общий | 1822 | ||||||||
| В88 |
| ||||||||||||
Две ветви R-V88, R-M18 и R-V35, встречаются почти исключительно на острове Сардиния .
Как видно из приведенной выше таблицы данных, R-V88 встречается в северном Камеруне и на западе Центральной Африки с очень высокой частотой, где его возникновение связывают с доисламским перемещением людей из Евразии . [64] [82]
R1b1b1 (R-M18)
R1b1b1 является подкладом R-V88, который определяется наличием маркера SNP M18. [7] Он был обнаружен только с низкой частотой в образцах из Сардинии [60] [83] и Ливана . [84]
R1b2 (R-PH155)
R1b2 встречается крайне редко и определяется наличием PH155. [36] Живые самцы, несущие субклады R-PH155, были обнаружены в Бахрейне , Бутане , Ладакхе , Таджикистане , Турции, Синьцзяне и Юньнани . ISOGG (2022) приводит две основные ветви: R-M335 (R1b2a) и R-PH200 (R1b2b).
Определяющий SNP R1b2a, M335, был впервые задокументирован в 2004 году, когда пример был обнаружен в Турции, хотя в то время он был классифицирован как R1b4. [34] Другие примеры R-M335 были зарегистрированы в образце хуэй из Юньнани , Китай [85] и в образце людей из Ладакха , Индия. [86]
Известные люди
Спытигнёв I, герцог Богемии , ДНК-тестирование его останков показало, что его Y-гаплогруппа была R1b. [87]
У династии Бурбонов , правивших во Франции, Испании и других европейских странах, есть гаплогруппа R1b1b. [88]
ДНК-тестирование нескольких мумий 18-й династии Египта выявило гаплогруппу R1b. Мумия Тутанхамона имела Y-гаплогруппу R1b и мтДНК-гаплогруппу K. Он унаследовал эту Y-гаплогруппу от своего отца, мумии KV55, которую многие считают Эхнатоном , и своего деда, Аменхотепа III , чья мумия была найдена погребенной в KV35 вместе с многочисленными родственниками. [89] [90]
Здоровье
Исследования показали, что гаплогруппа R1b может оказывать защитное действие на иммунную систему. [91] Однако более поздние исследования подтвердили, что Y-хромосома оказывает очень ограниченное влияние, например, на ишемическую болезнь сердца (ИБС), и что ранее предполагаемая связь между гаплогруппами Y-хромосомы и здоровьем далека от научного обоснования. [92]
Смотрите также
{kind=link}
Примечания
- ^ Флорес и др. (2005) обнаружили, что 20 из всех 146 протестированных мужчин (13,7%) — включая 20 из 45 протестированных мужчин из района Мертвого моря в Иордании — были положительными по M173 (R1) и отрицательными по обоим маркерам R1a SRY10831.2 и M17, а также P25 (который, как позже выяснилось, был ненадежным маркером для R1b1). Майрес и др. (2011) [41] указывают, что они в основном R-V88 (позже известный как R1b1b). Вуд и др. (2005) также сообщили о двух египетских случаях R1* (R-M173*), которые были отрицательными по SRY10831 (R1a1) и ненадежному маркеру R1b1 P25, из выборки из 1122 мужчин из африканских стран, включая 92 из Египта. Хассан и др. (2008) обнаружили столь же удивительное явление: 14 из 26 (54%) представителей народа фула из Судана имели M173+ и P25-.
- ^ Майрес и др. (2011): «Обнаружение V88 в Иране, Палестине и особенно в Мертвом море, Иордания (дополнительная таблица S4) дает представление о маршруте миграции обратно в Африку».
Ссылки
- ^ abc Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D и др. (июнь 2016 г.). «Генетическая история Европы ледникового периода». Nature . 534 (7606): 200– 5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. hdl :10211.3/198594. PMC 4943878 . PMID 27135931.
- ^ abcdef Хаак и др. 2015.
- ^ ab Аллентофт М.Э., Сикора М., Шегрен К.Г., Расмуссен С., Расмуссен М., Стендеруп Дж. и др. (июнь 2015 г.). «Популяционная геномика Евразии бронзового века». Природа . 522 (7555): 167–72 . Бибкод : 2015Natur.522..167A. дои : 10.1038/nature14507. PMID 26062507. S2CID 4399103.
- ^ ab Mathieson I, Lazaridis I, Rohland N, Mallick S, Patterson N, Roodenberg SA и др. (2015). «Восемь тысяч лет естественного отбора в Европе». bioRxiv : 016477. doi :10.1101/016477. S2CID 7866359.
- ^ Cassidy LM, Martiniano R, Murphy EM, Teasdale MD, Mallory J, Hartwell B, Bradley DG (январь 2016 г.). «Миграция в Ирландию в неолитический и бронзовый век и установление островного атлантического генома». Труды Национальной академии наук Соединенных Штатов Америки . 113 (2): 368–73 . Bibcode : 2016PNAS..113..368C. doi : 10.1073/pnas.1518445113 . PMC 4720318. PMID 26712024 .
- ^ Мартиниано Р., Кэссиди Л. М., О'Маолдуин Р., Маклафлин Р., Сильва Н. М., Манко Л. и др. (июль 2017 г.). «Популяционная геномика археологического перехода в Западной Иберии: исследование древней субструктуры с использованием методов импутации и гаплотипа». PLOS Genetics . 13 (7): e1006852. doi : 10.1371/journal.pgen.1006852 . PMC 5531429 . PMID 28749934.
- ^ abc Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и повышают разрешение древа гаплогруппы Y-хромосомы человека». Genome Research . 18 (5): 830– 8. doi :10.1101/gr.7172008. PMC 2336805 . PMID 18385274.
- ^ Mathieson 2018, Дополнительная таблица 1, строка 467.
- ^ Mathieson 2018, Дополнительная таблица 1, строки 251–272.
- ^ ab Джонс 2017.
- ^ Mathieson 2018, Дополнительная таблица 1, строки 205–245.
- ^ Mathieson 2018, Дополнительная таблица 1.
- ^ Матиесон 2018, стр. 2–3.
- ^ Гонсалес-Фортес и др. 2017, стр. 8–9.
- ^ Гонсалес-Фортес и др. 2017, с. 4, таблица 1, OC1_Meso.
- ^ Санчес-Куинто и др. 2019, Расширенный набор данных 1.3, OC1.
- ^ Mathieson 2018, Дополнительная таблица 1, строка 298, I4666.
- ^ Mathieson 2018, Дополнительная таблица 1, строка 153, I0124.
- ^ Mathieson 2018, Дополнительная таблица 1, строка 375, I0122.
- ^ Хаак и др. 2015, Дополнительная информация, стр. 26-27.
- ^ Хаак и др. 2015, Расширенная таблица данных 2, I0410.
- ^ Хаак и др. 2015, Дополнительная информация, стр. 44-45.
- ^ Mathieson 2018, Дополнительная таблица 1, I0410.
- ^ Mathieson 2018, Дополнительная таблица 1, строка 73, I2430.
- ^ Маркус и др. 2020, Дополнительные данные 1, Основная таблица, строка 25, MA89.
- ^ Mathieson 2018, Дополнительная таблица 1, строка 128, I0559.
- ^ de Barros Damgaard P, Martiniano R, Kamm J, Moreno-Mayar JV, Kroonen G, Peyrot M и др. (июнь 2018 г.). «Первые конные пастухи и влияние степных экспансий раннего бронзового века на Азию». Science . 360 (6396): eaar7711. doi :10.1126/science.aar7711. PMC 6748862 . PMID 29743352.
- ^ Хаак и др. 2015, стр. 5.
- ^ Karafet TM, Mendez FL, Sudoyo H, Lansing JS, Hammer MF (март 2015 г.). «Улучшенное филогенетическое разрешение и быстрая диверсификация гаплогруппы Y-хромосомы K-M526 в Юго-Восточной Азии». European Journal of Human Genetics . 23 (3): 369–73 . doi :10.1038/ejhg.2014.106. PMC 4326703. PMID 24896152 .
- ^ Сикора, Мартин; Питулько Владимир Владимирович; Соуза, Витор К.; Аллентофт, Мортен Э.; Виннер, Лассе; Расмуссен, Саймон; Маргарян, Ашот; де Баррос Дамгаард, Питер; де ла Фуэнте, Констанца; Рено, Габриэль; Ян, Мелинда А.; Фу, Цяомэй; Дюпанлуп, Изабель; Джампудакис, Константинос; Ногес-Браво, Давид (июнь 2019 г.). «История населения северо-восточной Сибири со времен плейстоцена». Природа . 570 (7760): 182–188 . Бибкод : 2019Natur.570..182S. doi : 10.1038/s41586-019-1279-z. hdl : 1887/3198847 . ISSN 1476-4687. PMID 31168093. S2CID 174809069.
- ^ Lacan M, Keyser C, Ricaut FX, Brucato N, Duranthon F, Guilaine J, et al. (Июнь 2011). «Древняя ДНК раскрывает мужскую диффузию по неолитическому средиземноморскому маршруту». Труды Национальной академии наук Соединенных Штатов Америки . 108 (24): 9788– 91. Bibcode : 2011PNAS..108.9788L. doi : 10.1073/pnas.1100723108 . PMC 3116412. PMID 21628562 .
- ^ Haak W, Balanovsky O, Sanchez JJ, Koshel S, Zaporozhchenko V, Adler CJ, et al. (Ноябрь 2010 г.). Penny D (ред.). «Древняя ДНК европейских фермеров раннего неолита раскрывает их ближневосточные связи». PLOS Biology . 8 (11): e1000536. doi : 10.1371/journal.pbio.1000536 . PMC 2976717. PMID 21085689 .
- ^ Olalde I, Brace S, Allentoft ME, Armit I, Kristiansen K, Booth T и др. (март 2018 г.). «Феномен Beaker и геномная трансформация северо-западной Европы». Nature . 555 (7695): 190– 196. Bibcode :2018Natur.555..190O. doi :10.1038/nature25738. PMC 5973796 . PMID 29466337.
- ^ abc Cinnioğlu C, King R, Kivisild T, Kalfoğlu E, Atasoy S, Cavalleri GL, et al. (январь 2004 г.). «Раскопки слоев гаплотипа Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127– 48. doi :10.1007/s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- ^ ab "ISOGG 2016 Y-ДНК Гаплогруппа R". isogg.org .
- ^ abc "ISOGG 2017 Y-ДНК Гаплогруппа R". isogg.org .
- ^ YПолный YTree v10.00.00
- ^ Филогенетическое дерево гаплогруппы R-V1636 на 23mofang
- ^ Гаплогруппа R на гаплодереве Y-ДНК на FTDNA
- ^ "R-V1636 YTree". www.yfull.com .
- ^ abcdefghijklm Майрес Н.М., Роотси С., Лин А.А., Ярве М., Кинг Р.Дж., Кутуев И. и др. (январь 2011 г.). «Эффект основателя основной гаплогруппы Y-хромосомы R1b голоцена в Центральной и Западной Европе». Европейский журнал генетики человека . 19 (1): 95–101 . doi :10.1038/ejhg.2010.146. ПМК 3039512 . ПМИД 20736979.
- ^ Grugni V, Battaglia V, Hooshiar Kashani B, Parolo S, Al-Zahery N, Achilli A и др. (18 июля 2012 г.). «Древние миграционные события на Ближнем Востоке: новые подсказки из вариации Y-хромосомы современных иранцев». PLOS ONE . 7 (7): e41252. Bibcode :2012PLoSO...741252G. doi : 10.1371/journal.pone.0041252 . PMC 3399854 . PMID 22815981.
- ^ Адамс SM, Кинг TE, Бош E, Джоблинг MA (май 2006 г.). «Дело о ненадежном SNP: повторяющаяся обратная мутация Y-хромосомного маркера P25 через генную конверсию». Forensic Science International . 159 (1): 14– 20. doi : 10.1016/j.forsciint.2005.06.003. hdl : 2381/443 . PMID 16026953.
- ^ abc Cruciani F, Тромбетта Б, Селлито Д, Массайя А, Дестро-Бисол Г, Уотсон Э и др. (июль 2010 г.). «Гаплогруппа R-V88 Y-хромосомы человека: отцовская генетическая запись транссахарских связей начала среднего голоцена и распространения чадских языков». Европейский журнал генетики человека . 18 (7): 800–7 . doi :10.1038/ejhg.2009.231. ПМЦ 2987365 . ПМИД 20051990.
- ^ Варзари А. (2006). История населения Днестровско-Карпатского региона: доказательства полиморфизма Alu-хромосомы и полиморфизмов Y-хромосомы (диссертация). Университет Людвига-Максимилиана в Мюнхене. doi :10.5282/edoc.5868.
- ^ ab Malyarchuk B, Derenko M, Denisova G, Maksimov A, Wozniak M, Grzybowski T, et al. (август 2011 г.). «Древние связи между сибиряками и коренными американцами, выявленные путем субтипирования гаплогруппы Y-хромосомы Q1a». Journal of Human Genetics . 56 (8): 583– 8. doi : 10.1038/jhg.2011.64 . PMID 21677663. S2CID 12015336.
- ^ Дулик М.С., Жаданов СИ, Осипова Л.П., Аскапули А., Гау Л., Гоккумен О. и др. (февраль 2012 г.). «Митохондриальная ДНК и вариации Y-хромосомы свидетельствуют о недавнем общем происхождении коренных американцев и коренных алтайцев». Американский журнал генетики человека . 90 (2): 229– 46. doi :10.1016/j.ajhg.2011.12.014. PMC 3276666. PMID 22281367 .
- ^ Behar DM, Yunusbayev B, Metspalu M, Metspalu E, Rosset S, Parik J, et al. (Июль 2010). «Структура еврейского народа на уровне генома». Nature . 466 (7303): 238– 42. Bibcode :2010Natur.466..238B. doi :10.1038/nature09103. PMID 20531471. S2CID 4307824.
- ^ Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, Edmonds CA, Chow CE и др. (февраль 2006 г.). «Полярность и временность распределений Y-хромосомы высокого разрешения в Индии выявляют как местные, так и экзогенные расширения и выявляют незначительное генетическое влияние скотоводов Центральной Азии». American Journal of Human Genetics . 78 (2): 202– 21. doi :10.1086/499411. PMC 1380230 . PMID 16400607.
- ^ аб Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента». ПЛОС ОДИН . 8 (10): е76748. Бибкод : 2013PLoSO...876748D. дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668.
- ^ Lippold S, Xu H, Ko A, Li M, Renaud G, Butthof A и др. (2014). «Отцовские и материнские демографические истории человека: выводы из последовательностей Y-хромосомы и мтДНК с высоким разрешением». Investigative Genetics . 5 : 13. doi : 10.1186/2041-2223-5-13 . PMC 4174254. PMID 25254093 .
- ^ Карафет ТМ, Осипова ЛП, Савина ОВ, Холлмарк Б, Хаммер МФ (ноябрь 2018 г.). «Сибирское генетическое разнообразие выявляет сложные истоки самодийских популяций». Американский журнал биологии человека . 30 (6): e23194. doi :10.1002/ajhb.23194. PMID 30408262. S2CID 53238849.
- ^ Аширбеков Е.Е., Ботбаев Д.М., Белкожаев А.М., Абайлдаев А.О., Неупокоева А.С., Мухатаев Ж.Э. и др. (2017). "Распределение гаплогрупп И-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей" [Распределение гаплогрупп Y-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей] (PDF) . Доклады Национальной академии наук Республики Казахстан (на русском языке). 6 (316): 85–95 . Архивировано из оригинала (PDF) 23 сентября 2021 г. Проверено 20 ноября 2021 г.
- ^ Шуху Л.И., Илихаму Н.И., Баке Р.А., Бупатима А.Б., Матюсуп Д.О. (2018). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica . 37 (1): 146–56 . doi :10.16359/j.cnki.cn11-1963/q.2017.0067.
- ^ R1b1a1b (R-M269) ранее была R1b1a1a2. С 2003 по 2005 год то, что сейчас называется R1b1a1b, обозначалось как R1b3. С 2005 по 2008 год это было R1b1c. С 2008 по 2011 год это было R1b1b2. С 2011 по 2018 год это было R1b1a1a2.
- ^ ab Balaresque P, Bowden GR, Adams SM, Leung HY, King TE, Rosser ZH и др. (январь 2010 г.). Penny D (ред.). "Преимущественно неолитическое происхождение европейских отцовских линий". PLOS Biology . 8 (1): e1000285. doi : 10.1371/journal.pbio.1000285 . PMC 2799514 . PMID 20087410.
- ^ Arredi B, Poloni ES, Tyler-Smith C (2007). "Заселение Европы". В Crawford MH (ред.). Антропологическая генетика: теория, методы и приложения . Кембридж, Великобритания: Cambridge University Press. стр. 394. ISBN 978-0-521-54697-3.
- ^ Cruciani F, Trombetta B, Antonelli C, Pascone R, Valesini G, Scalzi V и др. (июнь 2011 г.). «Сильная внутри- и межконтинентальная дифференциация, выявленная с помощью однонуклеотидных полиморфизмов Y-хромосомы M269, U106 и U152». Forensic Science International. Genetics . 5 (3): e49-52. doi :10.1016/j.fsigen.2010.07.006. hdl :11573/226727. PMID 20732840.
- ^ Груни, Виола; Равеан, Алессандро; Маттиоли, Франческа; Батталья, Винченца; Сала, Чинция; Тониоло, Даниэла; Ферретти, Лука; Гарделла, Рита; Ахилли, Алессандро; Оливьери, Анна; Торрони, Антонио; Пассарино, Джузеппе; Семино, Орнелла (февраль 2018 г.). «Реконструкция генетической истории итальянцев: новые идеи с точки зрения мужчины (Y-хромосомы)». Анналы биологии человека . 45 (1): 44–56 . doi :10.1080/03014460.2017.1409801. ISSN 1464-5033. PMID 29382284. S2CID 43501209.
- ^ ab Underhill PA, Shen P, Lin AA, Jin L, Passarino G, Yang WH и др. (ноябрь 2000 г.). «Изменение последовательности Y-хромосомы и история человеческих популяций». Nature Genetics . 26 (3): 358– 361. doi :10.1038/81685. PMID 11062480. S2CID 12893406.
- ^ Лобов АС (2009). Структура генофонда субпопуляций башкир (PDF) (диссертация на соискание ученой степени кандидата наук). Институт биохимии и генетики Уфимского научного центра РАН. Архивировано из оригинала (PDF) 2011-08-16.
- ^ Ордоньес AC, Фрегель Р., Трухильо-Медерос А, Эрвелла М, де-ла-Руа С, Арне-де-ла-Роса М (2017). «Генетические исследования доиспанского населения, захороненного в пещере Пунта-Асуль (Эль-Йерро, Канарские острова)». Журнал археологической науки . 78 : 20–28 . Бибкод :2017JArSc..78...20O. дои : 10.1016/j.jas.2016.11.004.
- ^ Robino C, Crobu F, Di Gaetano C, Bekada A, Benhamamouch S, Cerutti N и др. (май 2008 г.). «Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке алжирской популяции». International Journal of Legal Medicine . 122 (3): 251– 5. doi :10.1007/s00414-007-0203-5. PMID 17909833. S2CID 11556974.
- ^ abc Wood ET, Stover DA, Ehret C, Destro-Bisol G, Spedini G, McLeod H, et al. (Июль 2005 г.). «Контрастные закономерности вариации Y-хромосомы и мтДНК в Африке: доказательства демографических процессов, обусловленных полом». European Journal of Human Genetics . 13 (7): 867–76 . doi : 10.1038/sj.ejhg.5201408 . PMID 15856073. S2CID 20279122.
- ^ Епископосян Л, Худоян А, Арутюнян А (2006). «Генетическое тестирование гипотезы замены языка в Юго-Западной Азии». Иран и Кавказ . 10 (2): 191– 208. doi :10.1163/157338406780345899. JSTOR 4030922. S2CID 162345193.
- ^ Масиамо Хэй. «Гаплогруппа R1b (Y-ДНК)». Евпедия .
- ↑ Вадимовна Т.Н. (февраль 2015). Изменчивость Митохондриальной ДНК и Y-хромосомы в популяциях Волго-Уральского региона [ Вариации митохондриальной ДНК и Y-хромосомы у населения Волго-Уральского региона ] (PDF) (Диссертация по биологическим наукам) (на русском языке). Архивировано из оригинала (PDF) 2 апреля 2017 г.
- ^ Herrera KJ, Lowery RK, Hadden L, Calderon S, Chiou C, Yepiskoposyan L, et al. (март 2012 г.). «Неолитические патрилинейные сигналы указывают на то, что Армянское плато было повторно заселено земледельцами». European Journal of Human Genetics . 20 (3): 313– 20. doi :10.1038/ejhg.2011.192. PMC 3286660 . PMID 22085901.
- ^ Vanek D, Saskova L, Koch H (июнь 2009 г.). «Анализ родства и Y-хромосомы останков человека 7-го века: новая процедура извлечения и типирования ДНК для древнего материала». Хорватский медицинский журнал . 3. 50 (3): 286– 95. doi :10.3325/cmj.2009.50.286. PMC 2702742. PMID 19480023 .
- ^ D'Atanasio E, Trombetta B, Bonito M, Finocchio A, Di Vito G, Seghizzi M и др. (февраль 2018 г.). «Заселение последней Зеленой Сахары, выявленное с помощью высокопокрытия повторного секвенирования транссахарских патрилинейных линий». Genome Biology . 19 (1): 20. doi : 10.1186/s13059-018-1393-5 . PMC 5809971 . PMID 29433568.
- ^ Маркус Дж. Х., Пост С., Рингбауэр Х., Лай Л., Скитс Р., Сидоре С. и др. (февраль 2020 г.). «Генетическая история от среднего неолита до представления на средиземноморском острове Сардиния». Природные коммуникации . 11 (1): 939. Бибкод : 2020NatCo..11..939M. дои : 10.1038/s41467-020-14523-6. ПМК 7039977 . ПМИД 32094358.
- ^ ab Grugni, Viola; et al. (2019). «Анализ Y-хромосомы и фамилии для реконструкции прошлых популяционных структур: популяция Сардинии как тестовый случай». Международный журнал молекулярных наук . 20 (5763): 5763. doi : 10.3390/ijms20225763 . PMC 6888588. PMID 31744094 .
- ^ Аллентофт, ME (2024). "Популяционная геномика постледниковой Западной Евразии". Nature . 625 (7994): Дополнительная информация, стр. 48. Bibcode :2024Natur.625..301A. doi :10.1038/s41586-023-06865-0. PMC 10781627 . PMID 38200295.
- ^ Хабер, Марк; Меззавилла, Массимо; Бергстрем, Андерс; Прадо-Мартинес, Хавьер; Халласт, Пилле; Саиф-Али, Эр-Рияд; Аль-Хабори, Молхам; Дедусси, Джордж; Зеггини, Элефтерия; Блю-Смит, Джейсон; Уэллс, Р. Спенсер; Сюэ, Яли; Заллуа, Пьер А.; Тайлер-Смит, Крис (декабрь 2016 г.). «Генетическое разнообразие Чада раскрывает африканскую историю, отмеченную многочисленными голоценовыми евразийскими миграциями» (PDF) . Американский журнал генетики человека . 99 (6): 1316–1324 . doi :10.1016/j.ajhg.2016.10.012. ПМК 5142112 . PMID 27889059. S2CID 38169172.
- ^ Куличова, Ива; и др. (2017). «Внутренняя диверсификация не-южнее Сахары гаплогрупп в популяциях Сахеля и распространение скотоводства за пределами Сахары». Американский журнал биологической антропологии . 164 (2): 424– 434. doi :10.1002/ajpa.23285. PMID 28736914.
- ^ Висенте, Марио и др. (2019). «История популяции и генетическая адаптация кочевников фулани: выводы из данных по всему геному и признака устойчивости к лактазе». BMC Genomics . 20 (1): 915. doi : 10.1186/s12864-019-6296-7 . PMC 6888939. PMID 31791255 .
- ^ Симоеш, LG; и др. (2023). «Неолит северо-западной Африки, инициированный мигрантами из Иберии и Леванта». Nature . 618 (7965): 550– 556. Bibcode :2023Natur.618..550S. doi :10.1038/s41586-023-06166-6. PMC 10266975 . PMID 37286608.
- ^ Фрегель, Роза; Мендес, Фернандо Л.; Бокбот, Юсеф; Мартин-Сокас, Димас; Камалич-Массие, Мария Д.; Сантана, Джонатан; Моралес, Джейкоб; Авила-Аркос, Мария К.; Андерхилл, Питер А.; Шапиро, Бет; Войчик, Женевьева; Расмуссен, Мортен; Соарес, Андре ЭР; Капп, Джошуа; Сокелл, Александра (26 июня 2018 г.). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб как из Леванта, так и из Европы». Труды Национальной академии наук . 115 (26): 6774–6779 . Бибкод : 2018PNAS..115.6774F. doi : 10.1073/pnas.1800851115 . ISSN 0027-8424 . PMC 6042094. PMID 29895688.
- ^ Флорес С., Мака-Мейер Н., Ларруга Дж. М., Кабрера В. М., Карадшех Н., Гонсалес А. М. (2005). «Изоляты в коридоре миграций: высокоразрешающий анализ вариации Y-хромосомы в Иордании». Журнал генетики человека . 50 (9): 435–441 . doi : 10.1007/s10038-005-0274-4 . PMID 16142507. S2CID 6490283.
- ^ Хассан HY, Андерхилл PA, Кавалли-Сфорца LL, Ибрагим ME (ноябрь 2008 г.). «Изменение Y-хромосомы среди суданцев: ограниченный поток генов, соответствие языку, географии и истории». Американский журнал физической антропологии . 137 (3): 316– 323. doi :10.1002/ajpa.20876. PMID 18618658.
13/32
- ^ Шрайнер, Дэниел; Ротими, Чарльз Н. (декабрь 2018 г.). «Генетическая история Чада». Американский журнал физической антропологии . 167 (4): 804– 812. doi :10.1002/ajpa.23711. ISSN 0002-9483. PMC 6240361. PMID 30259956 .
- ^ Cruciani F, Santolamazza P, Shen P, Macaulay V, Moral P, Olckers A и др. (май 2002 г.). «Обратная миграция из Азии в Африку к югу от Сахары подтверждается высокоразрешающим анализом гаплотипов Y-хромосомы человека». American Journal of Human Genetics . 70 (5): 1197– 214. doi :10.1086/340257. PMC 447595 . PMID 11910562. , стр. 13–14
- ^ Contu D, Morelli L, Santoni F, Foster JW, Francalacci P, Cucca F (январь 2008 г.). "Доказательства на основе Y-хромосомы донеолитического происхождения генетически однородного, но разнообразного населения Сардинии: выводы для ассоциативного сканирования". PLOS ONE . 3 (1): e1430. Bibcode :2008PLoSO...3.1430C. doi : 10.1371/journal.pone.0001430 . PMC 2174525 . PMID 18183308.
- ^ Заллуа П.А., Сюэ Ю., Халифе Дж., Махул Н., Дебиан Л., Платт Д.Э. и др. (апрель 2008 г.). «Разнообразие Y-хромосом в Ливане структурировано недавними историческими событиями». Американский журнал генетики человека . 82 (4): 873–882 . doi :10.1016/j.ajhg.2008.01.020. ПМЦ 2427286 . ПМИД 18374297.
- ^ Zhong H, Shi H, Qi XB, Duan ZY, Tan PP, Jin L и др. (январь 2011 г.). «Расширенное исследование Y-хромосомы предполагает постледниковые миграции современных людей в Восточную Азию по северному маршруту». Молекулярная биология и эволюция . 28 (1): 717–27 . doi : 10.1093/molbev/msq247 . PMID 20837606.
- ^ Rowold DJ, Perez Benedico D, Garcia-Bertrand R, Chennakrishnaiah S, Alfonso-Sanchez MA, Gayden T, Herrera RJ (март 2016 г.). «Ладакх, Индия: земля высоких перевалов и генетической гетерогенности выявляет слияние миграций». European Journal of Human Genetics . 24 (3): 442– 9. doi :10.1038/ejhg.2015.80. PMC 4755386. PMID 25966630 .
- ^ "Odkud přišli Přemyslovci? Анализ ДНК был про vědce velkým překvapením" . dotyk.cz (на чешском языке) . Проверено 13 мая 2023 г.
- ^ Лармюзо, Маартен HD; Делорм, Филипп; Жермен, Патрик; Вандерхейден, Нэнси (9 октября 2013 г.). «Генетическая генеалогия раскрывает истинную гаплогруппу Y дома Бурбонов, противоречащую недавней идентификации предполагаемых останков двух французских королей». Eur J Hum Genet . 22 (5): 681– 687. doi :10.1038/ejhg.2013.211. PMC 3992573. PMID 24105374 .
- ^ Гад, Йехия (2020). «Материнская и отцовская родословная в семье царя Тутанхамона». Хранитель Древнего Египта: Эссе в честь Захи Хавасса. Чешский институт египтологии. С. 497–518 . ISBN 978-80-7308-979-5.
- ^ Гад, Йехия (2020). «Выводы из анализа древней ДНК египетских человеческих мумий: ключи к болезням и родству». Молекулярная генетика человека . 30 (R1): R24 – R28 . doi : 10.1093/hmg/ddaa223 . ISSN 0964-6906. PMID 33059357.
- ^ Маан А.А., Илс Дж., Акбаров А., Роуленд Дж., Сюй X, Джоблинг М.А. и др. (ноябрь 2017 г.). «Y-хромосома: основа мужского здоровья?». Европейский журнал генетики человека . 25 (11): 1181–1188 . doi :10.1038/ejhg.2017.128. ПМЦ 5643963 . ПМИД 28853720.
- ^ Тиммерс П., Уилсон Дж. Ф. (июль 2022 г.). «Ограниченное влияние вариации Y-хромосомы на ишемическую болезнь сердца и смертность в биобанке Великобритании — краткий отчет». Артериосклероз, тромбоз и сосудистая биология . 42 (9): 1198– 1206. doi : 10.1161 /ATVBAHA.122.317664. PMC 9394501. PMID 35861954.
Библиография
- Jones ER, Zarina G, Moiseyev V, Lightfoot E, Nigst PR, Manica A и др. (февраль 2017 г.). «Неолитический переход в Прибалтике не был обусловлен смешением с ранними европейскими земледельцами». Current Biology . 27 (4): 576– 582. Bibcode :2017CBio...27..576J. doi :10.1016/j.cub.2016.12.060. PMC 5321670 . PMID 28162894.
- González-Fortes G, Jones ER, Lightfoot E, Bonsall C, Lazar C, Grandal-d'Anglade A и др. (июнь 2017 г.). «Палеогеномные свидетельства многопоколенческого смешения неолитических земледельцев и мезолитических охотников-собирателей в бассейне Нижнего Дуная». Current Biology . 27 (12): 1801–1810.e10. Bibcode :2017CBio...27E1801G. doi :10.1016/j.cub.2017.05.023. PMC 5483232 . PMID 28552360.
- Haak W, Lazaridis I, Patterson N, Rohland N, Mallick S, Llamas B и др. (июнь 2015 г.). «Массовая миграция из степи стала источником индоевропейских языков в Европе». Nature . 522 (7555): 207– 211. arXiv : 1502.02783 . Bibcode :2015Natur.522..207H. doi :10.1038/nature14317. PMC 5048219 . PMID 25731166.
- Hollard C, Zvénigorosky V, Kovalev A, Kiryushin Y, Tishkin A, Lazaretov I, et al. (Сентябрь 2018). «Новые генетические доказательства сходства и разрывов между популяциями Сибири бронзового века». American Journal of Physical Anthropology . 167 (1): 97– 107. doi :10.1002/ajpa.23607. PMID 29900529. S2CID 205337212.
- Липсон М., Сеченьи-Надь А., Маллик С., Поса А., Стегмар Б., Кирл В. и др. (ноябрь 2017 г.). «Параллельные палеогеномные разрезы раскрывают сложную генетическую историю ранних европейских земледельцев». Природа . 551 (7680): 368–372 . Бибкод : 2017Natur.551..368L. дои : 10.1038/nature24476. ПМК 5973800 . ПМИД 29144465.
- Маркус Дж. Х., Пост С., Рингбауэр Х., Лай Л., Скитс Р., Сидоре С. и др. (февраль 2020 г.). «Генетическая история от среднего неолита до представления на средиземноморском острове Сардиния». Природные коммуникации . 11 (1): 939. Бибкод : 2020NatCo..11..939M. дои : 10.1038/s41467-020-14523-6. ПМК 7039977 . ПМИД 32094358.
- Мэтисон И., Алпаслан-Руденберг С., Пост С., Сечени-Надь А., Роланд Н., Маллик С. и др. (март 2018 г.). «Геномная история Юго-Восточной Европы». Природа . 555 (7695): 197–203 . Бибкод : 2018Natur.555..197M. дои : 10.1038/nature25778. ПМК 6091220 . ПМИД 29466330.
- Нарасимхан В.М., Паттерсон Н., Мурджани П., Роланд Н., Бернардос Р., Маллик С. и др. (сентябрь 2019 г.). «Формирование человеческих популяций в Южной и Центральной Азии». Наука . 365 (6457): eaat7487. doi : 10.1126/science.aat7487. ПМК 6822619 . ПМИД 31488661.
- Olalde I, Brace S, Allentoft ME, Armit I, Kristiansen K, Booth T и др. (март 2018 г.). «Феномен Beaker и геномная трансформация северо-западной Европы». Nature . 555 (7695): 190– 196. Bibcode :2018Natur.555..190O. doi :10.1038/nature25738. PMC 5973796 . PMID 29466337.
- Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; и др. (2015). «Массовая миграция из степи является источником индоевропейских языков в Европе». Природа . 522 (7555): 207–211 . arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H. bioRxiv 10.1101/013433 . дои : 10.1038/NATURE14317. ПМК 5048219 . ПМИД 25731166.
- Sánchez-Quinto F, Malmström H, Fraser M, Girdland-Flink L, Svensson EM, Simões LG и др. (май 2019 г.). «Мегалитические гробницы в западной и северной неолитической Европе были связаны с родственным обществом». Труды Национальной академии наук Соединенных Штатов Америки . 116 (19): 9469– 9474. Bibcode : 2019PNAS..116.9469S. doi : 10.1073/pnas.1818037116 . PMC 6511028. PMID 30988179.
- Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D и др. (июнь 2016 г.). «Генетическая история Европы ледникового периода». Nature . 534 (7606): 200– 5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. PMC 4943878 . PMID 27135931.
{{|url=https://www.dotyk.cz/magazin/premyslovci-dna-30000216.html}}
