Удильщик
| Удильщик Временной диапазон: | |
|---|---|
 | |
| Горбатый удильщик , Melanocetus johnsonii | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Анималия |
| Тип: | Хордовые |
| Сорт: | Актиноптеригии |
| Клад : | Перкоморфа |
| Заказ: | Лофиеобразные Гарман , 1899 |
| Типовой вид | |
| Рыболов-рыболов | |
{kind=link}
{kind=link}

(B) Cryptopsaras couesii , SL 34,5 мм
(C) Himantolophus appelii , SL 124 мм
(D) Diceratias trilobus , SL 86 мм
(E) Bufoceratias wedli , SL 96 мм
(F) Bufoceratias shaoi , SL 101 мм
(G) Melanocetus eustalus , SL 93 мм
(H) Lasiognathus amphirhamphus , SL 157 мм
(I) Thaumatichthys binghami , SL 83 мм
(J) Chaenophryne quasiramifera , SL 157 мм.
Рыба -удильщик — рыба из отряда костистых Lophiiformes ( / ˌ l ɒ f i ɪ ˈ f ɔːr m iː z / ). [1] Это костистые рыбы, названные так из-за их характерного способа хищничества , при котором модифицированный люминесцентный плавниковый луч (эска или иллиций) действует как приманка для других рыб. Свечение происходит от симбиотических бактерий , которые, как считается, приобретаются из морской воды, [2] [3] которые обитают в море и около него.
Некоторые удильщики отличаются экстремальным половым диморфизмом и половым симбиозом мелкого самца с гораздо более крупной самкой, что наблюдается в подотряде Ceratioidei , глубоководных удильщиках. У этих видов самцы могут быть на несколько порядков меньше самок. [4]
Рыбы-удильщики встречаются по всему миру. Некоторые из них пелагические (обитающие вдали от морского дна), в то время как другие бентосные (обитающие близко к морскому дну). Некоторые живут в глубоком море (например, Ceratiidae ), в то время как другие живут на континентальном шельфе , например, рыбы-лягушки и Lophiidae (рыба-чёрт или морской чёрт). Пелагические формы чаще всего сжаты с боков , тогда как бентосные формы часто чрезвычайно сжаты дорсовентрально (вдавлены), часто с большими устремленными вверх ртами. [ требуется цитата ]
Эволюция
Самые ранние окаменелости удильщиков были обнаружены в эоценовой формации Монте-Болка в Италии , и они уже демонстрируют значительную диверсификацию в современные семейства, составляющие отряд. [5] Учитывая это, а также их тесную связь с Tetraodontiformes , которые известны по меловым окаменелостям, они, вероятно, возникли в меловой период. [6] [7]
Филогенетическое исследование митохондриального генома 2010 года показало, что удильщики диверсифицировались в короткий период от начала до середины мелового периода, между 130 и 100 миллионами лет назад. [8] Более поздний препринт сокращает это время до позднего мелового периода , между 92 и 61 миллионом лет назад. [7] Другие исследования указывают на то, что удильщики возникли только вскоре после мел-палеогенового вымирания как часть массивной адаптивной радиации перкоморфов , хотя это противоречит обширному разнообразию, уже известному для этой группы к эоцену. [7] [9] Исследование 2024 года показало, что все подотряды удильщикообразных, скорее всего, отделились друг от друга в позднем мелу и палеоцене , но многочисленные семейства глубоководных удильщикообразных ( Ceratioidei ), а также их характерный половой паразитизм возникли в эоцене в результате быстрой радиации после палеоцен-эоценового термического максимума . [10]
Классификация
Рыбы-удильщики классифицируются в 5-м издании « Рыбы мира» , как указано ниже, на 5 подотрядов и 18 семейств. [11] [12] Следующие таксоны были организованы для демонстрации их эволюционных взаимоотношений. [4] [13]
- Подотряд Lophioidei Regan , 1912 г.
- Семейство Lophiidae Rafinesque , 1810 (Морские черти и морские уточки)
- Подотряд Antennarioidei Regan, 1912 г.
- Семейство Antennariidae Jarocki 1822 (Рыбы-лягушки)
- Семейство Tetrabrachiidae Regan, 1912 (Четырехбровые рыбы)
- Семейство Lophichthyidae Boeseman, 1964 (Лофихтиевые рыбы-лягушки)
- Семейство Brachionichthyidae Gill , 1863 (Рука-удильщик или бородавчатые удильщики)
- Подотряд Chaunacoidei Pietsch & Grobecker, 1987 г.
- Семейство Chaunacidae Gill, 1863 (Морские жабы)
- Подотряд Ogcocephaloidei Pietsch, 1984 г.
- Семейство Ogcocephalidae Gill, 1893 (Нетопыри)
- Подотряд Ceratioidei Regan, 1912
- Семейство Caulophrynidae Goode & Bean , 1896 (Веерообразные)
- Семейство Neoceratiidae Regan, 1926 (Колючие морские дьяволы)
- Семейство Melanocetidae Gill, 1878 (Черные морские дьяволы)
- Семейство Himantolophidae Gill, 1861 (Рыбы-футболы)
- Семейство Diceratiidae Regan & Trewavas , 1932 (Двойные удильщики)
- Семейство Oneirodidae Gill, 1878 (Мечтатели)
- Семейство Thaumatichthyidae Смит и Рэдклифф , 1912 (рыболовы-волчьи ловушки)
- Семейство Centrophrynidae Bertelsen , 1951 (Колючие морские черти)
- Семейство Ceratiidae Gill, 1861 (Бородавчатые морские черти)
- Семейство Gigantactinidae Boulenger , 1904 (Обыкновенные удильщики)
- Семейство Linophrynidae Regan, 1925 (Leftvents)
Ниже показаны взаимоотношения подотрядов внутри Lophiiformes, изложенные в работе Питча и Гробекера 1987 года « Рыбы-лягушки мира: систематика, зоогеография и поведенческая экология» . [8]
| Лофиеобразные |
| |||||||||||||||||||||||||||
{kind=link}

_(16059517767).jpg){kind=link}
_(16059517767).jpg/1280px-Hairy_frogfish_(Antennarius_striatus)_(16059517767).jpg)
В ходе филогенетических исследований было обнаружено, что как Lophiiformes, так и Tetraodontiformes гнездятся внутри Acanthuriformes и, таким образом, классифицируются как клады внутри этого таксона. [14]
Анатомия
Все удильщики плотоядны и, таким образом, приспособлены для захвата добычи. Цвет их варьируется от темно-серого до темно-коричневого, у глубоководных видов большие головы с огромными ртами в форме полумесяца, полными длинных, похожих на клыки зубов, наклоненных внутрь для эффективного захвата добычи. Их длина может варьироваться от 2 до 18 см (1–7 дюймов), а некоторые виды достигают 100 см (39 дюймов), [15] но эта вариация в значительной степени обусловлена половым диморфизмом, при этом самки намного крупнее самцов. [16] Рыбы-лягушки и другие мелководные виды удильщиков являются хищниками, нападающими из засады, и часто кажутся замаскированными под камни, губки или водоросли. [17]
У удильщиков есть лоскут, или иллициум, ближе к дистальному концу их тела на первом из двух спинных плавников, который простирается до морды и действует как механизм приманки, когда добыча приближается лицом к лицу. [18] Иллициум двигается вперед и назад пятью различными парами мышц: а именно, более короткими мышцами-выпрямителями и депрессорами, которые управляют движением подвздошной кости, а также мышцами-инклинаторами, протракторами и ретракторами, которые способствуют движению птеригиофора . [ 19]
В частности, если рассматривать Cryptopsaras couesii , то этот глубоководный цератиоидный удильщик имеет уникальную вращательную биомеханику в своей мускулатуре. Прочные мышцы ретрактора и протрактора движутся в извилистом узоре в противоположных направлениях по длине птеригиофора, который существует в глубоком продольном гребне вдоль черепа. [19] Кроме того, длинный и тонкий наклонитель глубоководного цератиоидного удильщика обеспечивает отчетливо широкий диапазон переднего и заднего движения, помогая движению приманочного аппарата, чтобы помочь в засаде на добычу. [19]
У большинства взрослых самок цератиоидных удильщиков есть люминесцентный орган, называемый эской, на кончике модифицированного спинного луча (иллициум или удочка; происходит от латинского ēsca , «приманка»). Предполагается, что этот орган служит для заманивания добычи в темных, глубоководных средах, но также служит для привлечения внимания самцов к самкам для облегчения спаривания.
Источником свечения являются симбиотические бактерии, которые обитают внутри и вокруг эски, заключенные в чашеобразный отражатель, содержащий кристаллы, вероятно, состоящие из гуанина . Удильщики используют эти симбиотические отношения с внеклеточными светящимися бактериями. [20] Нетипичные для светящихся симбионтов, которые живут вне клеток хозяина, бактерии, обнаруженные в приманках удильщиков, испытывают эволюционный сдвиг к меньшим и менее развитым геномам (геномная редукция), чему способствуют расширения транспозонов . [20] Только несколько видов светящихся симбионтов могут ассоциироваться с глубоководными удильщиками. [3] У некоторых видов бактерии, привлеченные в эску, не способны светиться независимо от удильщика, что позволяет предположить, что они развили симбиотические отношения, и бактерии не способны синтезировать все химические вещества, необходимые для свечения, самостоятельно. Они зависят от рыбы, чтобы компенсировать разницу. Хотя самки большинства семейств удильщиков обладают биолюминесценцией, существуют исключения, в том числе семейства Caulophrynidae и Neoceratiidae . [21]
Бактериальные симбионты не обнаруживаются на постоянном уровне на всех стадиях развития удильщиков или на разных глубинах океана. [22] Секвенирование личиночных организмов подотряда Ceratioidei показывает отсутствие бактериальных симбионтов, в то время как секвенирование взрослых удильщиков показывает более высокие уровни биолюминесцентных бактериальных симбионтов. [22] Это коррелирует с мезопелагическим регионом, имеющим самые высокие уровни симбионтных отношений в образцах удильщиков, поскольку именно там взрослые удильщики находятся большую часть своей жизни после личиночной стадии. [22] Электронная микроскопия этих бактерий у некоторых видов показывает, что они являются грамотрицательными палочками, у которых отсутствуют капсулы , споры или жгутики . Они имеют двухслойные клеточные стенки и мезосомы . Пора соединяет эску с морской водой, что позволяет удалять мертвые бактерии и клеточные отходы, а также позволяет pH и тоничности культуральной среды оставаться постоянными. Это, а также постоянная температура батипелагической зоны, в которой обитают эти рыбы, имеет решающее значение для долгосрочной жизнеспособности бактериальных культур. [23]
Световая железа всегда открыта наружу, поэтому возможно, что рыба приобретает бактерии из морской воды. Однако, похоже, что каждый вид использует свой собственный конкретный вид бактерий, и эти бактерии никогда не были обнаружены в морской воде. Хейгуд (1993) предположил, что эска выделяет бактерии во время нереста, и таким образом бактерии переносятся в икру. [23]
Некоторые данные показывают, что некоторые удильщики приобрели своих биолюминесцентных симбионтов из местной среды. Генетический материал бактерий-симбионтов обнаружен около удильщиков, что указывает на то, что удильщики и связанные с ними бактерии, скорее всего, не эволюционировали вместе, и бактерии проделывают сложный путь, чтобы попасть в хозяина. [3] В исследовании удильщиков-цератиоидов в Мексиканском заливе исследователи заметили, что подтвержденные связанные с хозяином биолюминесцентные микробы отсутствуют в личиночных особях и на протяжении всего развития хозяина. Цератиоиды, вероятно, приобрели своих биолюминесцентных симбионтов из морской воды. [2] Photobacteriumphosphoreum и члены клады kishitanii составляют основной или единственный биолюминесцентный симбионт нескольких семейств глубоководных светящихся рыб. [24]
Известно, что генетический состав бактерий-симбионтов претерпел изменения с тех пор, как они стали ассоциироваться со своим хозяином. [3] По сравнению с их свободноживущими родственниками, геномы симбионтов глубоководных удильщиков уменьшены в размере на 50%. Обнаружено сокращение путей синтеза аминокислот и способности использовать различные сахара. Тем не менее, гены, участвующие в хемотаксисе и подвижности, которые, как считается, полезны только вне хозяина, сохраняются в геноме. Геном симбионта содержит очень большое количество псевдогенов и демонстрирует массивное расширение мобильных элементов . Процесс редукции генома все еще продолжается в этих линиях симбионтов, и потеря гена может привести к зависимости от хозяина. [25]
У большинства видов широкий рот простирается по всему переднему краю головы, а полосы наклоненных внутрь зубов выстилают обе челюсти. Зубы могут быть вдавлены так, чтобы не создавать помех для объекта, скользящего к желудку, но не давать ему вырваться изо рта. [26] Рыба-удильщик способна растягивать как челюсть, так и желудок, поскольку ее кости тонкие и гибкие, до огромных размеров, что позволяет ей проглатывать добычу, которая в два раза больше ее тела. [27]
Поведение
Плавание и энергосбережение
В 2005 году недалеко от Монтерея, Калифорния , на глубине 1474 метра ROV в течение 24 минут снимал самку церациоидного удильщика рода Oneirodes . При приближении рыба быстро отступала, но в 74% видеоматериалов она дрейфовала пассивно, ориентируясь под любым углом. При движении вперед она плавала прерывисто со скоростью 0,24 длины тела в секунду, била грудными плавниками в фазе. Летаргическое поведение этого хищника, нападающего из засады, подходит для бедной энергией среды глубокого моря. [28]
Другое наблюдение in situ трех разных удильщиков-хлыстоносов показало необычное перевернутое поведение при плавании. Рыбы были замечены плавающими перевернутыми полностью неподвижно с иллициумом, свисающим вниз в виде небольшой дуги перед рыбой. Иллициум висел над небольшими видимыми норами. Было высказано предположение, что это попытка заманить добычу и пример низкоэнергетического оппортунистического поиска пищи и хищничества. Когда ROV приблизился к рыбе, она продемонстрировала взрывное плавание, все еще перевернутая. [29]
Челюсть и желудок удильщика могут расширяться, позволяя ему поглощать добычу, которая в два раза больше его самого. Из-за ограниченного количества пищи, доступной в среде обитания удильщика, эта адаптация позволяет удильщику запасать пищу, когда ее много. [30]
Хищничество
{kind=link}

Название «удильщик» происходит от характерного для вида метода хищничества. У удильщиков обычно есть по крайней мере одна длинная нить, вырастающая из середины их головы, называемая иллициумом. Иллициум — это отделенные и измененные первые три шипа переднего спинного плавника . У большинства видов удильщиков самая длинная нить — первая. Эта первая нить выступает над глазами рыбы и заканчивается нерегулярным наростом плоти (эской), и может двигаться во всех направлениях. Удильщики могут покачивать эской, чтобы она напоминала добычу, что заманивает добычу удильщика достаточно близко, чтобы удильщик мог поглотить ее целиком. [31] Некоторые глубоководные удильщики батипелагической зоны также излучают свет из своей эски , чтобы привлечь добычу. [32]
Поскольку удильщики являются оппортунистическими фуражировщиками, они демонстрируют диапазон предпочитаемой добычи с рыбами на крайних размерах спектра, в то же время демонстрируя повышенную избирательность к определенной добыче. Одно исследование, изучающее содержимое желудков нитеперых удильщиков у тихоокеанского побережья Центральной Америки, обнаружило, что эти рыбы в основном питались двумя категориями бентосной добычи: ракообразными и костистыми рыбами. Наиболее частой добычей были креветки-пандалиды . 52% исследованных желудков были пустыми, что подтверждает наблюдения о том, что удильщики являются потребителями низкого потребления энергии. [33]
Репродукция
.gif){kind=link}
.gif/1280px-Haplophryne_mollis_(female,_with_atrophied_male_attached).gif)
{kind=link}

Некоторые удильщики, такие как Ceratiidae, или морские дьяволы, используют необычный метод спаривания. [34] Поскольку отдельные особи локально редки, встречи также очень редки. Поэтому поиск пары проблематичен. Когда ученые впервые начали ловить цератиоидных удильщиков, они заметили, что все особи были самками. Эти особи были размером в несколько сантиметров, и почти у всех из них были прикреплены, по-видимому, паразиты . Оказалось, что эти «паразиты» были сильно диморфными самцами цератиоидов. Это указывает на то, что некоторые таксоны удильщиков используют полиандрическую систему спаривания. У некоторых видов удильщиков слияние самца и самки при размножении возможно из-за отсутствия ключей иммунной системы, которые позволяют антителам созревать и создавать рецепторы для Т-клеток. [35] Предполагается, что они выработали новые иммунные стратегии, которые компенсируют потерю функций В- и Т-лимфоцитов, обнаруженную в адаптивной иммунной системе . [36]
Некоторые цератиоиды полагаются на парабиотическое размножение. Свободноживущие самцы и непаразитированные самки этих видов никогда не имеют полностью развитых гонад. Таким образом, самцы никогда не созревают, не прикрепившись к самке, и умирают, если не могут ее найти. [4] При рождении самцы цератиоидов уже оснащены чрезвычайно хорошо развитыми обонятельными органами , которые обнаруживают запахи в воде. У самцов некоторых видов также развиваются большие, высокоспециализированные глаза, которые могут помочь в идентификации партнеров в темных условиях. Самцы цератиоидов значительно меньше самок удильщиков и могут испытывать трудности с поиском пищи в глубоком море. Кроме того, рост пищеварительных каналов некоторых самцов замедляется, что не позволяет им питаться. У некоторых таксонов челюсти никогда не подходят или неэффективны для захвата добычи. [37] Эти особенности означают, что самец должен быстро найти самку удильщика, чтобы предотвратить смерть. Чувствительные обонятельные органы помогают самцу обнаруживать феромоны , которые сигнализируют о близости самки удильщика.
Методы, которые используют удильщики для поиска партнеров, различаются. У некоторых видов крошечные глаза, которые не подходят для идентификации самок, в то время как у других недоразвитые ноздри, что делает маловероятным их эффективное нахождение самок по запаху. [4] Когда самец находит самку, он вгрызается в ее кожу и выделяет фермент , который переваривает кожу его рта и ее тела, сливая пару до уровня кровеносных сосудов. [36] Самец становится зависимым от самки-хозяина для выживания, получая питательные вещества через их общую кровеносную систему, и взамен предоставляет самке сперму. После слияния самцы увеличиваются в объеме и становятся намного крупнее по сравнению со свободно живущими самцами этого вида. Они живут и остаются репродуктивно функциональными до тех пор, пока жива самка, и могут принимать участие в нескольких нерестах. [4] Этот экстремальный половой диморфизм гарантирует, что когда самка готова к нересту, у нее немедленно есть партнер. [36] Несколько самцов могут быть объединены в одну самку, до восьми самцов у некоторых видов, хотя в некоторых таксонах, по-видимому, действует правило «один самец на самку». [4]
Симбиоз — не единственный метод размножения у удильщиков. На самом деле, многие семейства, включая Melanocetidae , Himantolophidae , Diceratiidae и Gigantactinidae , не демонстрируют никаких признаков симбиоза самцов. [37] Самки некоторых из этих видов содержат большие, развитые яичники, а свободноживущие самцы имеют большие яички, что позволяет предположить, что эти половозрелые особи могут метать икру во время временной половой связи, которая не предполагает слияния тканей. Самцы этих видов также имеют хорошо снабженные зубами челюсти, которые гораздо более эффективны при охоте, чем те, которые наблюдаются у симбиотических видов. [37]
Половой симбиоз может быть дополнительной стратегией у некоторых видов удильщиков. [4] У Oneirodidae самки, вынашивающие симбиотических самцов, были зарегистрированы у Leptacanthichthys и Bertella , а также у других, у которых еще не развились полностью функциональные гонады. [4] Одна из теорий предполагает, что самцы прикрепляются к самкам независимо от их собственного репродуктивного развития, если самка не достигла половой зрелости, но когда и самец, и самка достигают зрелости, они мечут икру, а затем разделяются. [4]
| Внешние видео | |
|---|---|
 Странный убийца глубин – YouTube Странный убийца глубин – YouTube | |
| Рыба-удильщик: оригинальный подход к глубоководной рыбалке – YouTube | |
| 3D-сканирование показывает огромную последнюю трапезу глубоководного удильщика – YouTube |
Одно из объяснений эволюции полового симбиоза заключается в том, что относительно низкая плотность самок в глубоководных средах оставляет мало возможностей для выбора партнера среди удильщиков. Самки остаются большими, чтобы обеспечить плодовитость , о чем свидетельствуют их большие яичники и икра. Самцы, как ожидается, будут уменьшаться, чтобы снизить метаболические затраты в средах с ограниченными ресурсами, и разовьют высокоспециализированные способности поиска самок. Если самцу удается найти самку, то симбиотическая привязанность в конечном итоге с большей вероятностью улучшит пожизненную пригодность по сравнению со свободной жизнью, особенно когда перспективы поиска будущих партнеров плохи. Дополнительным преимуществом симбиоза является то, что сперма самца может быть использована в нескольких оплодотворениях, поскольку он всегда остается доступным для самки для спаривания. Более высокая плотность встреч самцов и самок может коррелировать с видами, которые демонстрируют факультативный симбиоз или просто используют более традиционное временное контактное спаривание. [38]
Икра морского черта рода Lophius состоит из тонкого слоя прозрачного студенистого материала шириной 25 см (10 дюймов) и длиной более 10 м (33 футов). [39] Такой слой икры встречается редко у рыб. Икринки в этом слое располагаются в один слой, каждая в своей полости. Икра свободно плавает в море. Личинки свободно плавают и имеют брюшные плавники, вытянутые в нити. [26]
Угрозы
Виды Lophius, обитающие на северо-западе Европы, подвергаются интенсивному вылову и занесены в список ICES как «находящиеся вне безопасных биологических пределов». [40] Кроме того, известно, что удильщики иногда поднимаются на поверхность во время Эль-Ниньо , оставляя большие группы мертвых удильщиков плавающими на поверхности. [40]
В 2010 году Greenpeace International добавила американского удильщика ( Lophius americanus ), морского черта ( Lophius piscatorius ) и черноголового удильщика ( Lophius budegassa ) в свой Красный список морепродуктов — список рыб, которые обычно продаются по всему миру и с высокой вероятностью выловлены в результате неустойчивого рыболовства.Рыбы, занесенные в Красный список [41] [42]
Потребление человеком
Одно семейство, Lophiidae , представляет коммерческий интерес, его промыслы встречаются в Западной Европе, восточной части Северной Америки, Африке и Восточной Азии. В Европе и Северной Америке мясо хвоста рыбы рода Lophius , известной как морской черт или рыба-удав (Северная Америка), широко используется в кулинарии и часто сравнивается с хвостом омара по вкусу и текстуре.
В Африке наибольшие уловы зафиксированы в Намибии и Южно-Африканской Республике. [43] В Азии, особенно в Японии, печень морского черта, известная как анкимо , считается деликатесом. [44] Морской черт особенно активно потребляется в Южной Корее, где он является основным ингредиентом таких блюд, как агуджим .
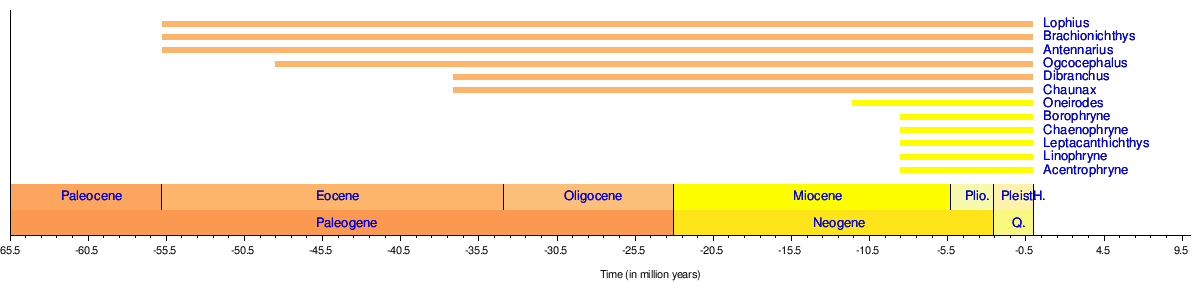
Хронология родов
В палеонтологической летописи удильщики представлены следующим образом: [45] [46] [47] [48]
Ссылки
- ^ Froese, Rainer ; Pauly, Daniel (ред.). "Order Lophiiformes". FishBase . Версия от февраля 2006 г.
- ^ ab Freed, Lindsay L.; Easson, Cole; Baker, Lydia J.; Fenolio, Dante; Sutton, Tracey T.; Khan, Yasmin; Blackwelder, Patricia; Hendry, Tory A.; Lopez, Jose V. (1 октября 2019 г.). «Характеристика микробиома и биолюминесцентных симбионтов на разных стадиях жизни цератиоидных удильщиков Мексиканского залива». FEMS Microbiology Ecology . 95 (10): fiz146. doi :10.1093/femsec/fiz146. ISSN 0168-6496. PMC 6778416 . PMID 31504465.
- ^ abcd Бейкер, Лидия Дж.; Фрид, Линдси Л.; Иссон, Коул Г.; Лопес, Хосе В.; Фенолио, Данте; Саттон, Трейси Т.; Найхолм, Спенсер В.; Хендри, Тори А. (1 октября 2019 г.). «Разнообразные глубоководные удильщики имеют генетически редуцированный светящийся симбионт, который приобретается из окружающей среды». eLife . 8 : e47606. doi : 10.7554/eLife.47606 . ISSN 2050-084X. PMC 6773444 . PMID 31571583.
- ^ abcdefghi Pietsch, Theodore W. (25 августа 2005 г.). «Диморфизм, паразитизм и пол снова: способы размножения среди глубоководных цератиоидных удильщиков (Teleostei: Lophiiformes)». Ichthyological Research . 52 (3): 207– 236. Bibcode : 2005IchtR..52..207P. doi : 10.1007/s10228-005-0286-2. S2CID 24768783.
- ^ Питч, Теодор В.; Карневале, Джорджио (2011). «Новый род и вид удильщикообразных (Teleostei: Lophiiformes: Lophiidae) из эоцена Монте-Болка, Италия». Copeia . 2011 (1): 64–71 . doi :10.1643/CI-10-080. ISSN 0045-8511.
- ^ Шане, Бруно; Гуинтард, Клод; Бетти, Эрик; Галлут, Кирилл; Деттаи, Агнес; Лекуантр, Гийом (1 сентября 2013 г.). «Доказательства тесной филогенетической связи между отрядами костистых костей Tetraodontiformes и Lophiiformes, основанные на анализе мягкой анатомии / Des donnees d'anatomie molle corroborent l'etroite Parente entre Tetraodontiformes et Lophiiformes». Цибиум . 37 (3): 179–199 .
- ^ abc Miller, Elizabeth Christina; Faucher, Rose; Hart, Pamela B.; Rincon-Sandoval, Melissa; Santaquiteria, Aintzane; White, William T.; Baldwin, Carole C.; Miya, Masaki; Betancur-R, Ricardo (30 октября 2023 г.), Phylogenomics reveals the deep ocean as a accelerator for evolutionary diversification in a fisherfishes, doi : 10.1101/2023.10.26.564281 , получено 21 декабря 2023 г.
- ^ ab Miya, M.; T. Pietsch ; J. Orr; R. Arnold; T. Satoh; A. Shedlock; H. Ho; M. Shimazaki; M. Yabe (2010). "Эволюционная история удильщикообразных (Teleostei: Lophiiformes): митогеномная перспектива". BMC Evolutionary Biology . 10 (1): 58. Bibcode : 2010BMCEE..10...58M. doi : 10.1186/1471-2148-10-58 . PMC 2836326. PMID 20178642 .
- ^ Альфаро, Майкл Э.; Фэрклот, Брант К.; Харрингтон, Ричард К.; Соренсон, Лори; Фридман, Мэтт; Такер, Кристин Э.; Оливерос, Карл Х.; Черны, Дэвид; Ниар, Томас Дж. (2018). «Взрывная диверсификация морских рыб на границе мелового и палеогенового периодов». Nature Ecology & Evolution . 2 (4): 688– 696. Bibcode : 2018NatEE...2..688A. doi : 10.1038/s41559-018-0494-6. ISSN 2397-334X. PMID 29531346.
- ^ Браунштейн, Чейз Д.; Цапфе, Катерина Л.; Лотт, Спенсер; Харрингтон, Ричард; Гезелайаг, Ава; Дорнбург, Алекс; Ниар, Томас Дж. (2024). «Синергические инновации способствовали распространению удильщиков в глубоком открытом океане». Current Biology . doi : 10.1016/j.cub.2024.04.066 . ISSN 0960-9822. PMID 38788708.
- ^ Нельсон, JS ; Гранде, TC; Уилсон, MVH (2016). Рыбы мира (5-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons . С. 508–518 . doi :10.1002/9781119174844. ISBN 978-1-118-34233-6. LCCN 2015037522. OCLC 951899884. ОЛ 25909650М.
- ^ Ричард ван дер Лаан; Уильям Н. Эшмейер и Рональд Фрике (2014). «Названия групп семейств современных рыб». Zootaxa . 3882 (2): 1– 230. doi : 10.11646/zootaxa.3882.1.1 . PMID 25543675.
- ^ Арнольд, Рэйчел Дж. (2014). Эволюционные связи загадочных удильщикообразных (Teleostei: Lophiiformes): может ли ядерная ДНК разрешить конфликтующие морфологические и митохондриальные филогении? (диссертация). Вашингтонский университет .
- ^ Near, Thomas J.; Thacker, Christine E. (2024). «Филогенетическая классификация современных и ископаемых лучеперых рыб (Actinopterygii)». Бюллетень Музея естественной истории Пибоди . 65 (1): 3– 302. doi : 10.3374/014.065.0101 .
- ^ "Anglerfish". National Geographic . 10 сентября 2010 г. Получено 28 февраля 2019 г.
- ^ "Идентификация рыб". fishbase.org . Получено 28 января 2019 г. .
- ^ Пембери Смит, Матильда QR; Ракстон, Грэм Д. (2020). «Камуфляж у хищников». Biological Reviews . 95 (5): 1325– 1340. doi : 10.1111/brv.12612. hdl : 10023/19948 . ISSN 1464-7931. PMID 32410297. S2CID 218649415.
- ^ Ясуги, Масаки; Хори, Мичио (июнь 2016 г.). «Преобладание параллельного и перекрестного хищничества у удильщиков». Морская экология . 37 (3): 576– 587. Bibcode : 2016MarEc..37..576Y. doi : 10.1111/maec.12309.
- ^ abc Симадзаки, Мицуоми; Накая, Казухиро (1 февраля 2004 г.). «Функциональная анатомия приманочного аппарата глубоководного цератиоидного удильщика Cryptopsaras couesii (Lophiiformes: Ceratiidae)». Ихтиологические исследования . 51 (1): 33–37 . Бибкод : 2004IchtR..51...33S. дои : 10.1007/s10228-003-0190-6. ISSN 1616-3915. S2CID 21508894.
- ^ ab Hendry, Tory A.; Freed, Lindsay L.; Fader, Dana; Fenolio, Dante; Sutton, Tracey T.; Lopez, Jose V. (5 июля 2018 г.). Moran, Nancy A. (ред.). «Продолжающееся транспозон-опосредованное сокращение генома у светящихся бактериальных симбионтов глубоководных цератиоидных удильщиков». mBio . 9 (3): e01033–18. doi :10.1128/mBio.01033-18. ISSN 2161-2129. PMC 6020299 . PMID 29946051.
- ^ Ludt, William B.; Clardy, Todd R. (март 2022 г.). «Первое обнаружение биофлуоресценции у глубоководного удильщика» . Journal of Fish Biology . 100 (3): 843– 846. Bibcode : 2022JFBio.100..843L. doi : 10.1111/jfb.14988. ISSN 0022-1112. PMID 34982469. S2CID 245670526.
- ^ abc Freed, Lindsay L.; Easson, Cole; Baker, Lydia J.; Fenolio, Dante; Sutton, Tracey T.; Khan, Yasmin; Blackwelder, Patricia; Hendry, Tory A.; Lopez, Jose V. (2019). "Характеристика микробиома и биолюминесцентных симбионтов на разных стадиях жизни цератиоидных удильщиков Мексиканского залива". FEMS Microbiology Ecology . 95 (10). doi :10.1093/femsec/fiz146. PMC 6778416 . PMID 31504465 . Получено 26 марта 2023 г. .
- ^ ab Munk, Ole; Hansen, Kjeld; Herring, Peter J. (2009). "О развитии и структуре светового органа эскал некоторых глубоководных удильщикообразных семейства меланоцитид (Pisces: Ceratioidei)". Журнал Морской биологической ассоциации Соединенного Королевства . 78 (4): 1321. doi :10.1017/S0025315400044520. ISSN 0025-3154. S2CID 84603596.
- ^ Хейгуд, Марго Г.; Дистель, Дэниел Л. (май 1993 г.). «Биолюминесцентные симбионты рыб-фонарей и глубоководных удильщиков образуют уникальные родословные, связанные с родом Vibrio». Nature . 363 (6425): 154– 156. Bibcode :1993Natur.363..154H. doi :10.1038/363154a0. ISSN 0028-0836. PMID 7683390. S2CID 4346611.
- ^ Хендри, Тори А.; Фрид, Линдси Л.; Фейдер, Дана; Фенолио, Данте; Саттон, Трейси Т.; Лопес, Хосе В. (26 июня 2018 г.). Моран, Нэнси А. (ред.). «Продолжающееся транспозон-опосредованное сокращение генома у светящихся бактериальных симбионтов глубоководных цератиоидных удильщиков». mBio . 9 (3): e01033–18, /mbio/9/3/mBio.01033–18.atom. doi :10.1128/mBio.01033-18. ISSN 2150-7511. PMC 6020299 . PMID 29946051.
- ^ ab Одно или несколько из предыдущих предложений включают текст из публикации, которая сейчас находится в общественном достоянии : Chisholm, Hugh , ed. (1911). "Angler". Encyclopaedia Britannica . Vol. 2 (11th ed.). Cambridge University Press. p. 15.
- ^ "Anglerfish". National Geographic . 10 сентября 2010 г. Получено 18 апреля 2018 г.
- ^ Лак, Даниэль Гарсия; Питч, Теодор В. (4 июня 2008 г.). «Наблюдения за глубоководным цератиоидным удильщиком рода Oneirodes (Lophiiformes: Oneirodidae)». Copeia . 2008 (2): 446– 451. doi :10.1643/CE-07-075. S2CID 55297852.
- ^ Мур, Джон А. (31 декабря 2001 г.). «Поведение перевернутого вниз головой удильщика (Teleostei: Ceratioidei: Gigantactinidae)». Copeia . 4. 2002 (4): 1144– 1146. doi :10.1643/0045-8511(2002)002[1144:udsbia]2.0.co;2. JSTOR 1448539. S2CID 85724627.
- ^ «Глубоководные удильщики — Глубоководные существа в море и небе».
- ^ Смит, Уильям Джон (2009). Поведение общения: этологический подход. Издательство Гарвардского университета. С. 381. ISBN 978-0-674-04379-4.
Другие полагаются на технику, принятую волком в овечьей шкуре, — они имитируют безобидные виды. ... Другие хищники даже имитируют добычу своей добычи: удильщикообразные (Lophiiformes) и аллигаторовые черепахи Macroclemys temmincki могут извиваться мясистыми выростами своих плавников или языков и привлекать мелких хищных рыб ко рту.
- ^ Пайпер, Росс (2007). Необыкновенные животные: энциклопедия любопытных и необычных животных , Greenwood Press .
- ^ Эспиноза, Марио; Инго Вертманн (2008). «Анализ содержимого желудка остроперого удильщика Lophiodes spilurus (Lophiiformes: Lophiidae), связанного с промыслом глубоководных креветок в центральной части Тихого океана Коста-Рики». Обзор тропической биологии . 4. 56 (4): 1959–70 . doi : 10.15517/rbt.v56i4.5772 . ПМИД 19419094 . Проверено 4 октября 2013 г.
- ^ Гори, Колм (23 марта 2018 г.). «Ученые были ошеломлены, запечатлев первую съемку спаривания странного удильщика». SiliconRepublic.com . Получено 23 марта 2018 г. .
- ^ Глубоководные удильщики выработали новый тип иммунной системы
- ^ abc Исаков, Ной (2022). «Гистосовместимость и воспроизводство: уроки удильщика». Life . 12 (1): 113. Bibcode :2022Life...12..113I. doi : 10.3390/life12010113 . ISSN 2075-1729. PMC 8780861 . PMID 35054506.
- ^ abc Pietsch, Theodore W. (8 марта 1972 г.). «Обзор семейства монотипных глубоководных удильщикообразных Centrophrynidae: таксономия, распространение и остеология». Copeia . 1972 (1): 17– 47. doi :10.2307/1442779. JSTOR 1442779.
- ^ Мия, Масаки; Питч, Теодор В; Орр, Джеймс В; Арнольд, Рэйчел Дж; Сато, Такаши П; Шедлок, Эндрю М; Хо, Сюань-Чин; Шимазаки, Мицуоми; Ябэ, Мамору; Нисида, Муцуми (1 января 2010 г.). "Эволюционная история удильщикообразных (Teleostei: Lophiiformes): митогеномная перспектива". BMC Evolutionary Biology . 10 (1): 58. Bibcode :2010BMCEE..10...58M. doi : 10.1186/1471-2148-10-58 . PMC 2836326 . PMID 20178642.
- ↑ Принс, EE 1891. Заметки о развитии рыбы-удильщика ( Lophius piscatorius ). Девятый ежегодный отчет Совета по рыболовству Шотландии (1890), Часть III: 343–348.
- ^ ab Clover, Charles (2004). The End of the Line: Как чрезмерный вылов рыбы меняет мир и то, что мы едим . Лондон: Ebury Press. ISBN 978-0-09-189780-2.
- ^ Красный список морепродуктов Гринпис
- ^ Питч, Теодор В. (2009). Океанические удильщики: необычайное разнообразие в глубоком море. Беркли: Издательство Калифорнийского университета. ISBN 978-0-520-94255-4. OCLC 1298208235.
- ^ Фаринья, AC; Азеведо, М; Ланда, Дж; Дуарте, Р.; Сампедро, П; Костас, Дж; Торрес, Массачусетс; Каньяс, Л. (октябрь 2008 г.). «Лофий в мире: синтез общих черт и жизненных стратегий». Журнал морских наук ICES . 65 (7): 1272–1280 . doi : 10.1093/icesjms/fsn140 .
- ^ "50 лучших блюд мира". CNN . CNN Travel. 12 июля 2017 г. Получено 27 марта 2023 г.
- ^ Сепкоски, Джек (2002). «Сводка родов ископаемых морских животных». Бюллетени американской палеонтологии . 364 : 560. Архивировано из оригинала 23 июля 2011 г. Получено 17 мая 2011 г.
- ^ Carnevale, Giorgio; Theodore W. Pietsch; Gary T. Takeuchi; Richard W. Huddleston (2008). "Fossil Ceratioid Anglerfishes (Teleostei: Lophiformes) from the Miocene of the Los Angeles Basin, California" (PDF) . Journal of Paleontology . 82 (5): 996– 1008. Bibcode :2008JPal...82..996C. doi :10.1666/07-113.1. S2CID 129926776. Архивировано из оригинала (PDF) 3 марта 2016 г. . Получено 27 октября 2022 г. .
- ^ Джорджио Карневале; Теордор В. Питч (12 июня 2009 г.). «Род глубоководных удильщиков Acentropryne (Teleostei, Ceratioidei, Linophrynidae) в миоцене Калифорнии» (PDF) . Журнал палеонтологии позвоночных . 29 (2): 372– 378. doi :10.1671/039.029.0232. ISSN 0272-4634. Wikidata Q114229338.
- ^ Назаркин, Михаил В.; Теодор В. Питч (2020). «Ископаемый сновидец рода Oneirodes (Lophiiformes: Ceratioidei) из миоцена острова Сахалин, Россия». Geological Magazine . 157 (8): 1378– 1382. Bibcode :2020GeoM..157.1378N. doi :10.1017/S0016756820000588. S2CID 225386060 . Получено 27 октября 2022 г. .
Дальнейшее чтение
- Андерсон, М. Эрик и Лесли, Робин В. 2001. Обзор глубоководных удильщикообразных (Lophiiformes: Ceratioidei) южной Африки. Ихтиологический бюллетень Института ихтиологии им. Дж. Л. Б. Смита; № 70. Институт ихтиологии им. Дж. Л. Б. Смита, Университет Родса
Внешние ссылки
{kind=link}
- Веб-проект «Дерево жизни»: Lophiiformes
- Видео (02:37) – Спаривание удильщиков на YouTube
- Лу, Д. Иммунная система удильщика позволяет ему сливаться со своим партнером. New Scientist 247, 19 (2020).
