Кератин для волос
Кератин волос — это тип кератина, который содержится в волосах и ногтях .
Функция
Возникнув из эмбрионального эпидермиса, волосяной фолликул развивается в одну из самых сложных структур в организме человека, состоящую из 7–8 отдельных участков ткани. [1] Основание волосяного фолликула содержит луковицу, в которой находятся дермальные фибробласты, известные как дермальный сосочек , имеющие решающее значение для морфогенеза и циклической активности волосяного фолликула. Вокруг этих клеток находится область матричных клеток, пролиферативный отсек волосяного фолликула, отвечающий за формирование различных отсеков фолликула (кроме ORS) и производство важнейших структурных элементов волоса — кератинов волос и связанных с ними белков, известных как KAP. [1]
Кератин — это важный волокнистый белок , встречающийся у животных, образующий такие жесткие структуры, как волосы, перья, ногти и рога. Он классифицируется на основе происхождения ткани и содержания серы: мягкие кератины содержат меньше серы, в то время как твердые кератины, встречающиеся в волосах и когтях, содержат больше серы, что создает более прочную структуру. [2] Кератины бывают двух типов — кислые Типа I и нейтрально-основные Типа II, далее подразделяющиеся на Тип I a и b, и Тип II a и b. Начальным этапом формирования кератина является выравнивание полипептидов кератина типа I и типа II для создания гетеродимера, который затем агрегируется в структурные единицы более высокого порядка. [2] Подобно другим белкам субъединиц промежуточных филаментов , существует распространенная вторичная структура: хорошо сохранившийся центральный альфа-спиральный домен, состоящий из четырех спирально-спиральных сегментов вместе с неспиральными конечными доменами, которые различаются по последовательностям и длине [14]. Недавние открытия показывают, что взаимодействие между кислыми и основными мягкими кератинами начинается с создания гетеродимера. Этот гетеродимер состоит из кислого и основного мономерного кератина. Два из этих гетеродимеров затем объединяются, образуя тетрамер, который впоследствии полимеризуется, что приводит к образованию конечной 10-нанометровой нитевидной структуры. [3]
Стабильность
Благодаря своей роли структурных стабилизаторов в эпителиальных клетках, кератиновые филаменты привлекли значительный интерес в биологии, эмбриологии, патологии и дерматологии. Эта фундаментальная функция цитоскелета выходит за рамки отдельных клеток. Обычно кератиновые филаменты интегрированы в десмосомы (см. рис. 1b, d) и гемидесмосомы, способствуя не только межклеточной стабильности, но и прикреплению к базальной мембране и соединительной ткани в пределах конкретного эпителия. [4] В нестратифицированном (простом) эпителии внутренних органов, испытывающем минимальную механическую нагрузку, только несколько типов кератина образуют редко распределенные филаменты в цитоплазме. Однако более значительное число типов кератина участвует в промежуточном филаментном цитоскелетном каркасе плоского эпителия, который становится более заметным в ороговевших стратифицированных эпителиях, таких как эпидермис, покрывающий внешнюю поверхность тела. Здесь кератины многочисленны и плотно упакованы, образуя тонофиламенты. [4]
Заживление ран
В последнее время внимание было привлечено к замечательным возможностям заживления ран и превосходной биосовместимости кератина, полученного из человеческих волос. Хотя рекомбинантные кератиновые белки, полученные с помощью технологии рекомбинантной ДНК, обладают более высокой чистотой по сравнению с извлеченным кератином, их ранозаживляющие свойства остаются неясными. Два рекомбинантных трихоцитарных кератина — человеческий кератин волос I типа 37 и человеческий кератин волос II типа 81 — были экспрессированы с использованием системы бактериальной экспрессии и впоследствии образовали рекомбинантные кератиновые наночастицы (RKNP) посредством ультразвуковой дисперсии. [5] Было обнаружено, что RKNP значительно усиливают пролиферацию и миграцию клеток в лабораторных условиях. Более того, при нанесении на дермальные раны in vivo, RKNP способствовали улучшению заживления ран, что приводило к усиленной эпителизации, васкуляризации, отложению коллагена и ремоделированию. Важно, что тесты на биосовместимость in vivo не показали признаков системной токсичности. RKNP имеют потенциал как многообещающий подход к ускорению заживления ран и предлагают новые пути для разработки биоматериалов на основе кератина. [5]
Уменьшение кровотечения
Исследования эффективности гемостаза in vivo проводились с использованием крысиных моделей пункции печени и повреждения бедренной артерии. Для обеих моделей K37 и K81 (10 мг) наносились на раневые области. В модели пункции печени время кровотечения значительно уменьшалось с рекомбинантным K37 (приблизительно 38 с) и K81 (приблизительно 40 с) по сравнению с одним только носителем (приблизительно 170 с, p < .01), при этом заметно уменьшалась общая потеря крови (p < .01). [6] Кроме того, в модели повреждения бедренной артерии рекомбинантные кератиновые белки значительно уменьшали время кровотечения по сравнению с контрольной группой (приблизительно 50 с против 270 с). Примечательно, что K37 и K81 продемонстрировали более сильные гемостатические эффекты, чем извлеченные кератины (приблизительно 80 с) при лечении повреждения печени крыс. Кроме того, рекомбинантные кератиновые белки продемонстрировали надежную способность способствовать формированию фибринового сгустка в месте повреждения, эффективно останавливая кровотечение. Следовательно, рекомбинантные кератины человеческих волос предлагают потенциал для разработки новых гемостатических продуктов на основе кератиновых биоматериалов. [6]
Типы
Существует два типа кератина для волос:
- кислый тип I кератин волос
- кератин волос типа I 1 , KRT31
- кератин волос типа I 2 , KRT32
- кератин волос типа I 3A , KRT33A
- кератин волос типа I 3B , KRT33B
- кератин волос типа I 4 , KRT34
- кератин волос типа I 5 , KRT35
- кератин волос 6 типа I , KRT36
- кератин волос типа I 7 , KRT37
- кератин волос типа I 8 , KRT38
- основной кератин волос типа II
- кератин волос II типа 1 , KRT81
- кератин волос II типа 2 , KRT82
- кератин волос II типа 3 , KRT83
- кератин волос II типа 4 , KRT84
- кератин волос II типа 5 , KRT85
- кератин волос 2 типа 6 , KRT86
Ассоциированные белки
{kind=link}
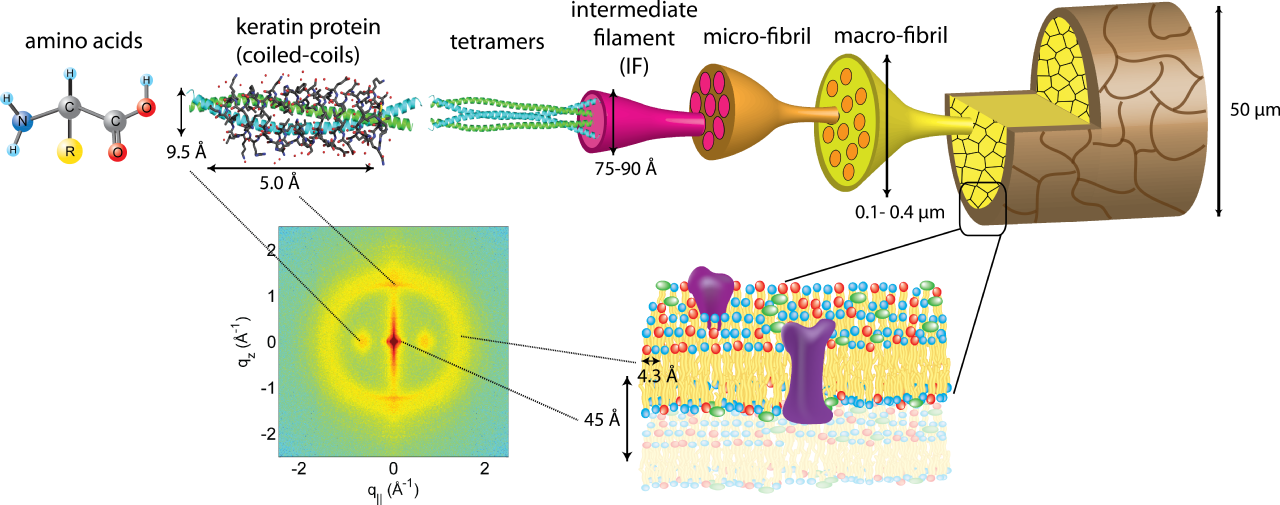
Стержень волоса в основном состоит из кератинов волос и связанных с ними белков (KRTAP). KRTAP являются продуктами различных семейств генов, возникших в результате событий дупликации генов в их эволюционной истории. Эти гены, как правило, небольшие, состоящие из одного экзона длиной менее 1000 пар оснований. За последнее десятилетие у млекопитающих, включая людей, было идентифицировано множество генов KRTAP. Они подразделяются на три группы в зависимости от их аминокислотного состава: высокосернистые (с <30 мол.% цистеина), сверхвысокосернистые (>30 мол.% цистеина) и высокоглициново-тирозиновые. [7] Кератины волос образуют промежуточные нити (KIF) внутри трихоцитов, специализированных клеток, которые способствуют формированию волос. По мере того, как эти клетки перемещаются вверх в коре, KIF агрегируют, окруженные пространством, называемым матриксом. KRTAP, также известные как KAP, являются значительной частью этой матрицы между KIF. Предполагается, что KRTAP играют роль в создании сшитой сети с KIF, способствуя формированию жесткого стержня волоса. [7]
Экспрессия генов
Во время роста волос, когда клетки фолликулярной луковицы быстро трансформируются в кортикальные или кутикулярные кератиноциты волос, на уровне транскрипции активируются приблизительно 50-100 генов кератина. [8] Однако этот сложный процесс можно упростить до нескольких высокосохраняемых семейств генов. В кортикальных кератиноцитах очевидны различные паттерны экспрессии генов кератина, что указывает на наличие различных иерархических процессов транскрипции среди различных типов клеток. Исследование областей промотора гена кератина выявляет консервативные мотивы последовательности, которые могут управлять этими специфичными для клеток признаками. [8] Более того, посредством изоляции родственных генов кератина кутикулы овец и человека были обнаружены консервативные мотивы ДНК и паттерны экспрессии во время дифференциации клеток кутикулы. Кроме того, экспрессия генов IF фолликула овечьей шерсти и высокосернистого кератина у трансгенных мышей предполагает, что регуляторные элементы ДНК и белки, связанные с генами кератина волос, поддерживают функциональную консервацию у разных видов млекопитающих. [8]
Клиническое значение
Рак молочной железы
Кератин представляет собой большое мультигенное семейство, известное как цитокератины. Эти цитокератины дифференциально экспрессируются в различных типах эпителия и были тщательно изучены в качестве маркеров рака молочной железы. Они подразделяются на кислые цитокератины типа I и основные-нейтральные цитокератины типа II. [9] Промежуточная сеть филаментов формируется необходимым спариванием равных количеств кератинов типа I и типа II. Хотя кератины волос, такие как KRT81, типичны для жестко кератинизированных структур, таких как волосы и ногти, считается, что они служат структурными белками, специфичными для этих органов, без экспрессии в других местах, таких как молочная железа.
KRT81, кератин волос II типа, является основным белком волос, экспрессируемым в корковом слое волоса. Интересно, что, несмотря на то, что он обычно ассоциируется со структурами волос, экспрессия KRT81 наблюдалась в клеточной линии рака молочной железы человека SKBR3 и метастатических лимфатических узлах карцином молочной железы, но не в нормальных эпителиальных клетках молочной железы. Более того, было обнаружено, что экспрессируемый KRT81 представляет собой 5′-укороченную изоформу (ΔHb1), при этом полноразмерный белок не экспрессируется. [9] Однако точная функция этой укороченной формы в клетках рака молочной железы остается неясной.
Анализ вестерн-блоттинга выявил наличие полного 55-кДа KRT81 в различных линиях клеток рака молочной железы человека (MCF7, SKBR3, MDA-MB-231), нормальных эпителиальных клетках молочной железы человека (HMEC) и неопухолевых клетках (MCF10A). [9] Обратная транскрипция-полимеразная цепная реакция подтвердила экспрессию полноразмерного KRT81, включая его 5'-область, в клетках молочной железы. Иммуногистохимические и иммунофлуоресцентные исследования обнаружили KRT81 в цитоплазме. Кроме того, в клетках MDA-MB231 с нокдауном KRT81 зимография продемонстрировала снижение активности MMP9, в то время как анализы царапин и инвазии продемонстрировали снижение миграции клеток и возможностей инвазии. [ 9] Это представляет собой первое доказательство полной экспрессии KRT81 как в нормальных эпителиальных клетках молочной железы, так и в клетках рака молочной железы. Кроме того, полученные данные свидетельствуют о том, что KRT81 играет роль в миграции и инвазии клеток рака молочной железы.
Ссылки
- ^ ab Rogers MA, Langbein L, Praetzel-Wunder S, Winter H, Schweizer J (январь 2006 г.). "Протеины, ассоциированные с кератином волос человека (KAP)". International Review of Cytology . 251. Academic Press: 209– 263. doi :10.1016/S0074-7696(06)51006-X. ISBN 9780123646552. PMID 16939781 . Получено 28.11.2023 .
- ^ ab Cruz CF, Azoia NG, Matamá T, Cavaco-Paulo A (август 2017 г.). «Взаимодействие пептидов и белков в кератинах человеческих волос». International Journal of Biological Macromolecules . 101 : 805– 814. doi : 10.1016/j.ijbiomac.2017.03.052. hdl : 1822/56408 . PMID 28315768.
- ^ Yu J, Yu DW, Checkla DM, Freedberg IM, Bertolino AP (июль 1993 г.). «Кератины человеческих волос». Журнал исследовательской дерматологии . 101 (1 Suppl): 56S – 59S . doi :10.1111/1523-1747.ep12362635. PMID 7686952.
- ^ ab Moll R, Divo M, Langbein L (июнь 2008 г.). «Человеческие кератины: биология и патология». Histochemistry and Cell Biology . 129 (6): 705– 733. doi :10.1007/s00418-008-0435-6. PMC 2386534. PMID 18461349 .
- ^ ab Gao F, Li W, Deng J, Kan J, Guo T, Wang B, Hao S (май 2019 г.). «Рекомбинантные наночастицы кератина человеческих волос ускоряют заживление дермальных ран». ACS Applied Materials & Interfaces . 11 (20): 18681– 18690. doi : 10.1021/acsami.9b01725. PMID 31038908. S2CID 141367865.
- ^ ab Guo T, Li W, Wang J, Luo T, Lou D, Wang B, Hao S (2018). «Рекомбинантные белки кератина человеческих волос для остановки кровотечения». Искусственные клетки, наномедицина и биотехнология . 46 (sup2): 456– 461. doi : 10.1080/21691401.2018.1459633 . PMID 29621887. S2CID 4620386.
- ^ ab Fujikawa H, Fujimoto A, Farooq M, Ito M, Shimomura Y (июль 2012 г.). «Характеристика семейства генов белка 2, ассоциированного с кератином волос человека (KRTAP2)». Журнал исследовательской дерматологии . 132 (7): 1806–1813 . doi : 10.1038/jid.2012.73 . PMID 22495175.
- ^ abc Пауэлл BC, Нески А, Роджерс GE (декабрь 1991 г.). «Регуляция экспрессии генов кератина при дифференцировке волосяных фолликулов». Анналы Нью-Йоркской академии наук . 642 : 1–20 . doi :10.1111/j.1749-6632.1991.tb24376.x. PMID 1725577. S2CID 33504645.
- ^ abcd Nanashima N, Horie K, Yamada T, Shimizu T, Tsuchida S (май 2017). «Кератин волос KRT81 экспрессируется в нормальных клетках и клетках рака молочной железы и способствует их инвазивности». Oncology Reports . 37 (5): 2964–2970. doi : 10.3892 /or.2017.5564 . PMID 28405679.
Внешние ссылки
- Кератины,+специфичные для волос в Национальной медицинской библиотеке США, медицинские предметные рубрики (MeSH)
- "Кератин волос". 31 мая 2006 г. Получено 30 марта 2017 г.
