Синдром острова

Синдром острова описывает различия в морфологии , экологии , физиологии и поведении островных видов по сравнению с их континентальными аналогами. Эти различия развиваются из-за различного экологического давления, влияющего на островные виды, включая малочисленность крупных хищников и травоядных, а также постоянно мягкий климат . [2] [3]
Экологические движущие факторы
- Сокращение хищничества . Островные экосистемы не могут поддерживать достаточную биомассу добычи, чтобы вместить крупных хищников. Это в значительной степени избавляет виды добычи от риска хищничества, [2] что в основном устраняет давление отбора на морфологии, экологию и поведение, которые помогают избегать крупных хищников.
- Сокращение биоразнообразия . Островные экосистемы, как правило, включают большие популяции ограниченного числа видов (состояние, называемое компенсацией плотности ), поэтому они демонстрируют низкое биоразнообразие . Это приводит к снижению межвидовой конкуренции и повышению внутривидовой конкуренции . [2]
- Снижение полового отбора . У островных видов также наблюдается снижение полового отбора , что особенно заметно у птиц, которые теряют свое половое диморфное оперение, используемое в сексуальных демонстрациях. [4]
- Уменьшение разнообразия паразитов . Наконец, в островных экосистемах наблюдается уменьшение разнообразия паразитов [5] , что снижает уровень отбора, действующего на гены, связанные с иммунитетом.
Особенности островного синдрома у животных
Размер тела

Межвидовая конкуренция между континентальными видами приводит к расхождению размеров тела, так что виды могут избегать высоких уровней конкуренции, занимая отдельные ниши . Снижение межвидовой конкуренции между островными видами снижает это давление отбора для видов, занимающих отдельные ниши. [6] В результате, существует меньшее разнообразие в размерах тела островных видов. Обычно мелкие млекопитающие увеличиваются в размерах (например, фосса является более крупным островным родственником мангуста ) , в то время как обычно крупные млекопитающие уменьшаются в размерах (например, малагасийские бегемоты являются более мелкими островными родственниками континентальных бегемотов ). Это примеры островного гигантизма и островной карликовости соответственно. Этот наблюдаемый эффект называется правилом Фостера . Напротив, птицы и рептилии, как правило, демонстрируют островной гигантизм, примером которого являются моа , казуар и комодский варан .
Хотя гигантские черепахи Галапагосских островов и Сейшельских островов ( галапагосская черепаха и Aldabrachelys соответственно) иногда приводятся в качестве примеров островного гигантизма [2] , в настоящее время считается, что они представляют собой последние оставшиеся популяции исторически широко распространенных гигантских черепах. Останки черепах аналогичного или большего размера были найдены в Австралии ( Meiolania ), Южной Азии ( Megalochelys ), Мадагаскаре ( Aldabrachelys ), Северной Америке [7] ( Hesperotestudo ) и Южной Америке [8] ( Chelonoidis ). Считается, что нынешние гигантские черепахи сохранились только на нескольких отдаленных архипелагах, поскольку люди прибыли туда относительно поздно и не сильно предшествовали им, что позволяет предположить, что эти популяции черепах были менее подвержены чрезмерной эксплуатации .
Передвижение
Поскольку островные виды добычи испытывают меньший риск нападения хищников, они часто теряют или уменьшают морфологии, используемые для уклонения от хищников. Например, крылья долгоносиков , пастушков и голубей стали настолько редуцированными у островных видов, что многие из них утратили способность летать. [2] Это произошло у нескольких бескилевых , включая киви и казуара, а также у додо и какапо после вторжения в островные среды обитания. Вымершие моа Новой Зеландии демонстрируют самый экстремальный известный пример редукции островных крыльев; нет никаких костных свидетельств даже рудиментарных крыльев , а плечевой пояс редуцирован до скапулокоракоидеума, который не смог бы нести переднюю конечность, поскольку у него отсутствует суставная ямка . [9] Таким образом, это единственная известная птица, полностью утратившая свои крылья после перехода к островному образу жизни. Потеря способности к полету позволяет птицам исключить затраты на поддержание крупных мышц, обеспечивающих полет, таких как грудные мышцы , и позволяет скелету стать тяжелее и прочнее. [10] Островные популяции сипух имеют более короткие крылья, что представляет собой переходную стадию, на которой их способность к полету снижается. [11]
- Киви Окарито — небольшая нелетающая птица семейства бескилевых , обитающая на Южном острове Новой Зеландии.
- Скелет вымершего моа из Новой Зеландии рядом с Ричардом Оуэном . Моа демонстрировал островной гигантизм , небольшой размер мозга [9] и полную потерю крыльев, включая элемент стилоподия . [9]
- Южный казуар — нелетающая бескилевая птица , обитающая в Индонезии , Новой Гвинее и северо-восточной Австралии [1]



![Скелет вымершего моа из Новой Зеландии рядом с Ричардом Оуэном. Моа демонстрировали островной гигантизм, небольшой размер мозга[9] и полную потерю крыльев, включая элемент стилоподия[9].](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bb/Dinornis1387.jpg/1280px-Dinornis1387.jpg)
Адаптивная окраска
Из-за сниженного полового отбора островных видов, они, как правило, демонстрируют сниженную половую окраску, чтобы сохранить энергию, которая для этого требуется. Кроме того, низкое биоразнообразие островных экосистем делает распознавание видов менее важным, поэтому видоспецифическая окраска находится под меньшим отбором. [5] В результате островные виды птиц часто демонстрируют более тусклое, полово-мономорфное оперение. [5]
Несколько островных видов приобретают повышенную окраску меланина . Самцы белокрылых расписных малюров, живущих на материковой части Австралии, демонстрируют синее брачное оперение, тогда как два островных подвида ( Malurus leucopterus leucopterus с острова Дирка Хартога и Malurus leucopterus edouardi с острова Барроу ) демонстрируют черное брачное оперение. [12] Подвид каштановобрюхого монарха, эндемичного для Соломоновых островов , Monarcha castaneiventris obscurior , демонстрирует полиморфизм в цвете оперения: некоторые птицы черные с каштановым животом, в то время как другие полностью меланические. Частота меланического фенотипа увеличивается на более мелких островах, даже если учитывать относительную близость островов. [13]
Репродукция
Высокий уровень внутривидовой конкуренции между потомками приводит к отбору наиболее приспособленных особей. В результате островные родители, как правило, производят меньше потомства, так что каждое потомство получает больший родительский вклад , максимизируя свою приспособленность . [2] Ящерицы, эндемичные для островных экосистем, откладывают меньшие кладки, которые дают большее потомство по сравнению с континентальными ящерицами аналогичного размера. Из-за более высокой частоты откладывания яиц у островных ящериц, континентальные и островные ящерицы производили потомство с сопоставимой скоростью. [14]
Размер мозга

Гипотеза дорогой ткани предполагает, что ткани с высокой метаболической потребностью, такие как мозг, будут уменьшаться, если они дают мало селективного преимущества и, таким образом, не способствуют увеличению потребления пищи. Малочисленность крупных хищников означает, что островные виды могут позволить себе стать медленнее и менее бдительными, не страдая от значительно возросшего риска нападения хищников. Таким образом, уменьшение относительного размера мозга часто наблюдается у островных видов, поскольку это снижает базальную скорость метаболизма [15] без увеличения риска нападения хищников. Например, эндокраниальный объем вымершего мадагаскарского карликового бегемота на 30% меньше, чем у континентального предка такого же размера. [16] Аналогичным образом, ранний человек , Homo floresiensis , имел мозг такого же размера, как и у значительно более ранних особей австралопитеков с материковой Африки [17] [18], и в 3,4 раза меньше, чем у Homo sapiens , который эволюционировал позже (см. Эволюция размера человеческого мозга ).
пойкилотермия
Из-за низкого риска нападения хищников островные виды добычи могут позволить себе снизить базальный уровень метаболизма , приняв пойкилотермию без значительного увеличения риска нападения хищников. В результате пойкилотермия гораздо более распространена среди островных видов. [10]
Поведение
Из-за отсутствия хищников островные виды, как правило, становятся более послушными и менее территориальными, чем их континентальные собратья (иногда это называют островной прирученностью ). [2] [19] Оленьи мыши , певчие воробьи и бронзовые анолисы имеют меньшие территории с большим перекрытием по сравнению с их материковыми сородичами. Они также более терпимы к незваным гостям. Фолклендские лисицы и таммарские валлаби утратили врожденный страх перед крупными хищниками, включая людей. [2]
У паразитов
Нематодный паразит Heligmosomoides polygyrus подвергся расширению ниши (путем вторжения в новые виды хозяев) и сокращению генетического разнообразия после вторжения в экосистемы семи островов западного Средиземноморья . Потеря генетического разнообразия была связана с расстоянием между современной популяцией и материковым происхождением. [20]
В растениях

Структура растения
Высота растений и площадь листьев следуют образцу островных млекопитающих, при этом мелкие виды становятся крупнее, а крупные виды — мельче в островных популяциях. [2] [6] Это может быть связано с уменьшением межвидовой конкуренции, что снижает экологический стимул для растений занимать отдельные ниши. Из-за сокращения биомассы крупных травоядных некоторые островные растения теряют защитные шипы и колючки, а также уменьшают количество производимых защитных химических веществ . Маловероятность островных пожаров также приводит к потере огнестойкости коры, плодов и шишек. Островная древесность, эволюционный переход от травянистости к древесности , является очень распространенным явлением среди островных флор. [21]
Размножение и расселение
Из-за отсутствия специализированных опылителей на отдаленных островах, островные растения часто используют маленькие, неброско окрашенные и легкодоступные цветы, чтобы привлечь ряд альтернативных видов опылителей. Самоопыление также чаще используется островными видами растений, поскольку пыльце не нужно путешествовать так далеко, чтобы достичь восприимчивой семяпочки или рыльца . Семена проявляют островной гигантизм , становясь преимущественно больше, чем семена с материка, что, как полагают, снижает смертность в море во время рассеивания . [2] [6]
Последствия островного синдрома для сохранения природы
Снижение риска хищничества в островных экосистемах привело к потере нескольких адаптаций и поведенческих моделей, которые позволяют избегать или препятствовать нападению хищников. Это делает островные виды особенно уязвимыми для эксплуатации чужеродными видами . Например, когда люди впервые завезли на остров Маврикий собак, свиней, кошек, крыс и макак, питающихся крабами, в XVII веке, они разграбили гнезда додо и усилили межвидовую конкуренцию за ограниченные пищевые ресурсы. [22] Это в конечном итоге привело к вымиранию додо. Ограниченные ресурсы в островных экосистемах также уязвимы для чрезмерной эксплуатации, если они не управляются устойчиво .
Остров Хацег


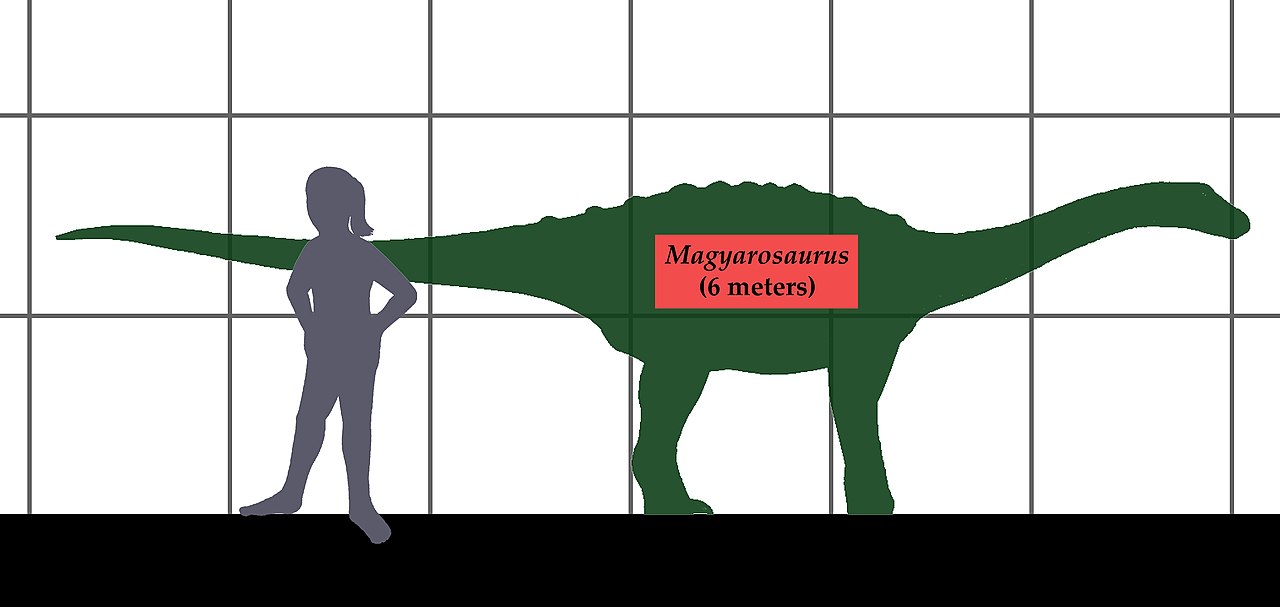
Остров Хацег был крупным прибрежным островом в море Тетис в позднем меловом периоде [23] и часто называется «Островом карликовых динозавров» из-за обширных ископаемых свидетельств того, что его местные динозавры демонстрировали островную карликовость . Местный титанозавр острова , Magyarosaurus dacus , имел массу тела всего 900 килограммов (2000 фунтов) [24] [25], в то время как материковые титанозавры , такие как Patagotitan, могли достигать 69 тонн (76 тонн). Птерозавр Hatzegopteryx занял позицию главного хищника острова Хацег в отсутствие каких-либо гиперхищных динозавров и, вероятно, охотился на молодых карликовых динозавров или даже взрослых особей более мелких видов. На основании его крепкой челюсти и шейных позвонков, Hatzegopteryx , как полагают, охотился аналогично современным аистам, нападая на добычу, которая слишком велика, чтобы проглотить ее целиком. [26] По оценкам, размах его крыльев достигал 10–12 метров (33–39 футов), что делает его одним из крупнейших птерозавров , когда-либо живших. Таким образом, он является ярким примером островного гигантизма , в данном случае для заполнения в противном случае пустой ниши высшего хищника . Balaur bondoc изначально был классифицирован как динозавр дромеозаврида на основе его втягивающихся когтей на пальцах ног. Его передние конечности казались слишком короткими и коренастыми, чтобы он был базальным avialan , однако филогенетический анализ позже подтвердил, что Balaur действительно был базальным членом Avialae , клады, которая включает современных птиц . Его конечности явно не были способны к активному полету, поэтому балаур является еще одним примером вторичной утраты способности к полету после вторжения на островную нишу, подобно бескилевым птицам , а также вымершим моа и додо (см. Островное снижение способности к полету).
Синдром обратного острова
Термин «синдром обратного острова» (RIS) был впервые использован Паскуале Райей в 2010 году для описания различий в морфологии , экологии , физиологии и поведении, наблюдаемых у островных видов, когда плотность популяции либо низкая, либо колеблется. [27] Это приводит к более сильному естественному отбору и более слабому внутривидовому отбору, что приводит к различным фенотипам по сравнению со стандартным островным синдромом.
RIS был впервые описан в популяции итальянской настенной ящерицы , эндемичной для острова Ликоза , где непредсказуемые условия окружающей среды и сильно колеблющаяся плотность популяции отобрали агрессивное поведение и повышенные репродуктивные усилия. [28] Самцы ящериц демонстрируют повышенные уровни α-МСГ по сравнению с материковыми популяциями, что увеличивает базальную скорость метаболизма, усиливает иммунные реакции, [29] производит более темную синюю окраску и повышает уровень 5α-дигидротестостерона . [28] Последний улучшает репродуктивный успех самцов, увеличивая вероятность победы в сексуальных конфликтах над самками и повышая качество спермы. [28] Самки производят такое же количество яиц, как и материковые популяции, но яйца островных самок значительно тяжелее, что отражает повышенные репродуктивные усилия. Непредсказуемые условия приводят к высоким показателям смертности, поэтому взрослые особи вкладывают больше усилий в текущие выводки, поскольку они с меньшей вероятностью выживут, чтобы произвести последующие выводки, т. е. существует низкий уровень межвыводковых конфликтов .
- Островной подвид итальянской настенной ящерицы Podarcis siculus klemmeri имеет более тёмную синюю окраску.
- Материковый фенотип итальянской настенной ящерицы с более светлой зелено-коричневой окраской


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Ссылки
- ^ ab Clements, James (2007). Список птиц мира Клементса (6-е изд.). Итака, Нью-Йорк: Cornell University Press. ISBN 978-0-8014-4501-9.
- ^ abcdefghijk Baeckens, Simon; Van Damme, Raoul (20 апреля 2020 г.). «Островной синдром». Current Biology . 30 (8): R329–R339. Bibcode : 2020CBio...30.R338B. doi : 10.1016/j.cub.2020.03.029 . PMID 32315628.
- ^ Гаврилиди, Иоанна; Де Мистер, Жиль; Ван Дамм, Рауль; Бэккенс, Саймон (2022). «Как вести себя, когда выброшен на берег: поведенческий компонент синдрома острова остается недостаточно изученным». Biology Letters . 18 (4): 20220030. doi : 10.1098/rsbl.2022.0030 . PMC 9039784. PMID 35440235 .
- ^ Гриффит, СК (2000). «Высокая точность на островах: сравнительное исследование внепарного отцовства у воробьиных птиц». Поведенческая экология . 11 (3): 265–273. doi : 10.1093/beheco/11.3.265 .
- ^ abc Bliard, L.; Paquet, M.; Robert, A.; Dufour, P.; Renoult, JP; Grégoire, A.; Crochet, P.; Covas, R.; Doutrelant, C. (22 апреля 2020 г.). «Изучение связи между расслабленным хищничеством и окраской птиц на островах». Biology Letters . 16 (4). doi :10.1098/rsbl.2020.0002. PMC 7211456 . PMID 32315593.
- ^ abc Biddick, M.; Hendriks, A.; Burns, KC (19 августа 2019 г.). «Растения подчиняются (и не подчиняются) правилу острова». Труды Национальной академии наук Соединенных Штатов Америки . 116 (36): 17632–17634. Bibcode : 2019PNAS..11617632B. doi : 10.1073/pnas.1907424116 . PMC 6731657. PMID 31427521 .
- ^ Хансен, Деннис М.; Донлан, К. Джош; Гриффитс, Кристин Дж.; Кэмпбелл, Карл Дж. (22 июня 2010 г.). «Экологическая история и скрытый потенциал сохранения: большие и гигантские черепахи как модель для замены таксонов» (PDF) . Ecography . 33 (2): 272–284. Bibcode :2010Ecogr..33..272H. doi :10.1111/j.1600-0587.2010.06305.x.
- ^ Cione, AL; Tonni, EP; Soibelzon, L. (2003). «The Broken Zig-Zag: Late Cenozoic large infant and tortoise bleach in South America» (PDF) . Rev. Mus. Argentino Cienc. Nat . NS 5 (1): 1–19. doi : 10.22179/REVMACN.5.26 . ISSN 1514-5158. Архивировано из оригинала (PDF) 2011-07-06.
- ^ abc Worthy, TH; Scofield, RP (2012). «Достижения XXI века в области знаний о биологии моа (Aves: Dinornithiformes): новый морфологический анализ и пересмотренные диагнозы моа». Новозеландский журнал зоологии . 39 (2): 87–153. doi :10.1080/03014223.2012.665060. S2CID 83768608.
- ^ ab McNab, BK (октябрь 1994 г.). «Сохранение энергии и эволюция нелетающей способности у птиц». The American Naturalist . 144 (4): 628–648. doi :10.1086/285697. JSTOR 2462941. S2CID 86511951.
- ^ Рулен, А.; Саламин, Н. (19 апреля 2010 г.). «Ограниченность и эволюция меланизма, полового дихроматизма и размера тела у распространенной по всему миру сипухи». Журнал эволюционной биологии . 23 (5): 925–934. doi : 10.1111/j.1420-9101.2010.01961.x . PMID 20298444. S2CID 20356466.
- ^ Уолш, Дженнифер; Кампанья, Леонардо; Фини, Уильям Э.; Кинг, Хасинта; Вебстер, Майкл С. (5 февраля 2021 г.). «Закономерности генетической дивергенции и демографическая история проливают свет на динамику популяций остров-материк и эволюцию меланического оперения у белокрылого расписного малюра». Международный журнал органической эволюции . 75 (6): 1348–1360. doi : 10.1111/evo.14185. PMID 33543771. S2CID 231820197.
- ^ Uy, J. Albert C.; Vargas-Castro, Luis E. (22 июля 2015 г.). «Размер острова предсказывает частоту встречаемости меланических птиц у полиморфной по цвету мухоловки Monarcha castaneiventris на Соломоновых островах». The Auk: Ornithological Advances . 132 (4): 787–794. doi : 10.1642/AUK-14-284.1 .
- ^ Новосолов, Мария; Райя, Паскуале; Мейри, Шай (22 августа 2012 г.). «Островной синдром у ящериц». Глобальная экология и биогеография . 22 (2): 184–191. doi :10.1111/j.1466-8238.2012.00791.x.
- ^ Herculano-Houzel, Suzana (1 марта 2011 г.). «Масштабирование метаболизма мозга с фиксированным энергетическим бюджетом на нейрон: последствия для нейронной активности, пластичности и эволюции». PLOS ONE . 6 (3): e17514. Bibcode :2011PLoSO...617514H. doi : 10.1371/journal.pone.0017514 . PMC 3046985 . PMID 21390261.
- ^ Уэстон, Элеанор М.; Листер, Адриан М. (7 мая 2009 г.). «Островная карликовость у бегемотов и модель уменьшения размера мозга у Homo floresiensis». Nature . 459 (7243): 85–88. Bibcode :2009Natur.459...85W. doi :10.1038/nature07922. PMC 2679980 . PMID 19424156.
- ^ Brown, P.; Sutikna, T.; Morwood, MJ; Soejono, RP; Jatmiko; Saptomo, E. Wayhu; Due, Rokus Awe (28 октября 2004 г.). «Новый мелкотелый гоминин из позднего плейстоцена Флореса, Индонезия». Nature . 431 (7012): 1055–1061. Bibcode :2004Natur.431.1055B. doi :10.1038/nature02999. PMID 15514638. S2CID 26441.
- ^ Фальк, Дин; Хильдеболт, Чарльз; Смит, Кирк; Морвуд, MJ; Сутикна, Томас; Браун, Питер; Джатмико; Саптомо, Э. Вайху; Брансден, Барри; Прайор, Фред (8 апреля 2005 г.). «Мозг LB1, Homo floresiensis». Наука . 308 (5719): 242–245. Бибкод : 2005Sci...308..242F. дои : 10.1126/science.1109727. PMID 15749690. S2CID 43166136.
- ^ Гаврилиди, Иоанна; Де Мистер, Жиль; Ван Дамм, Рауль; Бэккенс, Саймон (2022). «Как вести себя, когда выброшен на берег: поведенческий компонент синдрома острова остается недостаточно изученным». Biology Letters . 18 (4): 20220030. doi : 10.1098/rsbl.2022.0030 . PMC 9039784. PMID 35440235 .
- ^ Nieberding, C.; Morand, S.; Libois, R.; Michaux, JR (25 мая 2006 г.). «Паразиты и островной синдром: колонизация западных средиземноморских островов Heligmosomoides polygyrus (Dujardin, 1845)». Журнал биогеографии . 33 (7): 1212–1222. Bibcode : 2006JBiog..33.1212N. doi : 10.1111/j.1365-2699.2006.01503.x. hdl : 2268/78063 . S2CID 83545722.
- ^ Эволюция островной лесистости - PNAS
- ^ Хьюм, Дж. П.; Уолтерс, М. (2012). Вымершие птицы. Лондон: A & C Black. С. 134–136. ISBN 978-1-4081-5725-1.
- ^ Бентон, М. Дж., Чики, З., Григореску, Д., Редельсторфф, Р., Сандер, П. М., Стайн, К. и Вайсхампль, Д. Б. (2010). «Динозавры и правление острова: карликовые динозавры с острова Хацег. Архивировано 10 июля 2011 г. в Wayback Machine » Палеогеография, палеоклиматология, палеоэкология , 293 (3-4): 438–454.
- ^ Carballido, JL; Pol, D.; Otero, A.; Cerda, IA; Salgado, L.; Garrido, AC; Ramezani, J.; Cúneo, NR; Krause, JM (2017). «Новый гигантский титанозавр проливает свет на эволюцию массы тела среди динозавров-завропод». Труды Королевского общества B: Биологические науки . 284 (1860): 20171219. doi :10.1098/rspb.2017.1219. PMC 5563814. PMID 28794222 .
- ^ Stein, K.; Csiki, Z.; Rogers, KC; Weishampel, DB; Redelstorff, R.; Carballido, JL; Sander, PM (30 апреля 2010 г.). «Маленький размер тела и экстремальное ремоделирование кортикальной кости указывают на филетическую карликовость у Magyarosaurus dacus (Sauropoda: Titanosauria)». Труды Национальной академии наук . 107 (20): 9258–9263. Bibcode : 2010PNAS..107.9258S. doi : 10.1073/pnas.1000781107 . PMC 2889090. PMID 20435913 .
- ^ Виттон, М. П.; Наиш, Д. (2015). «Аждархидные птерозавры: имитаторы пеликанов, питающихся водой, или «наземные охотники»?». Acta Palaeontologica Polonica . 60 (3). doi : 10.4202/app.00005.2013 .
- ^ Райя, Паскуале; Гуарино, Фабио М; Турано, Миммо; Полезе, Джанлука; Риппа, Даниэла; Каротенуто, Франческо; Монти, Дарья М; Карди, Мануэла; Фульджоне, Доменико (20 сентября 2010 г.). «Спандрел голубая ящерица и островной синдром». Эволюционная биология BMC . 10 (289): 289. Бибкод : 2010BMCEE..10..289R. дои : 10.1186/1471-2148-10-289 . ПМЦ 2949876 . ПМИД 20854657.
- ^ abc Монти, Дарья Мария; Райя, Паскуале; Вроонен, Джессика; Маселли, Валерия; Ван Дамм, Рауль; Фульджионе, Доменико (январь 2013 г.). «Физиологические изменения в популяции островных ящериц подтверждают синдром обратного острова». Биологический журнал Линнеевского общества . 108 (1): 144–150. doi : 10.1111/j.1095-8312.2012.02019.x .
- ^ Baeckens, Simon; Van Damme, Raoul (2018). «Иммунокомпетентность и паразитарная инвазия в меланистической и нормально окрашенной популяции ящериц-лацертид, Podarcis siculus». Amphibia-Reptilia . 39 (4): 471–478. doi : 10.1163/15685381-20181008. hdl : 10067/1550590151162165141 . ISSN 0173-5373. S2CID 52245927.
Смотрите также
