Мадьярозавр
| Мадьярозавр Временной диапазон: Маастрихт , | |
|---|---|
 | |
| Плечевая кость, Музей естественной истории Девы | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Анималия |
| Тип: | Хордовые |
| Клад : | Динозавры |
| Клад : | Ящеротазовые |
| Клад : | † Зауроподоморфы |
| Клад : | † Зауроподы |
| Клад : | † Макронария |
| Клад : | † Титанозавры |
| Клад : | † Литостротия |
| Род: | † Магьярозавр фон Хюне , 1932 г. |
| Типовой вид | |
| † Титанозавр дакус | |
| Разновидность | |
| |
| Синонимы [1] | |
| |
{kind=link}
Magyarosaurus (« венгерский ящер») — род карликовых динозавров - завропод из позднего мелового периода (ранний-поздний маастрихт ) в Румынии . Это один из самых маленьких известных взрослых завропод, длиной всего 6 м (20 футов) и массой тела 750–1000 кг (1650–2200 фунтов). Тип и единственный определенный вид — Magyarosaurus dacus . В исследовании 2005 годабыло обнаружено, что он является близким родственником Rapetosaurus в семействе Saltasauridae в кладе завропод Titanosauria . [2]
Открытие
{kind=link}

Останки, принадлежащие по крайней мере десяти особям, были извлечены из региона Хунедоара ( формация Сынпетру ) в районе, который во время их открытия был Венгрией, но теперь является западной Румынией. Первоначально они были названы Titanosaurus dacus , видовое название отсылало к дакам (которые жили в этом месте около 2000 лет назад), бароном Нопчей в 1915 году. [3] Нопчей собирал окаменелости в этом районе с 1895 года. Позднее вид был переименован в Magyarosaurus dacus Фридрихом фон Хюне в 1932 году. [4] фон Хюне в 1932 году также назвал два других вида: M. hungaricus и M. transsylvanicus . Более крупный, более редкий M. hungaricus может представлять собой отдельный таксон . [5]
Голотип , BMNH R.3861a , состоит из набора позвонков. Было найдено множество других костей, в основном хвостовые позвонки, а также спинные и элементы аппендикулярного скелета. Остатки черепов неизвестны. Было обнаружено 14 ископаемых яиц, которые были приписаны Magyarosaurus . [6]
{kind=link}

Палеонтологические исследования проводились в Рапа Роше около Себеша , на юго-западной стороне Трансильванского бассейна. Исследования были начаты в 1969 году . Кости динозавров были обнаружены в более ранних исследованиях. На основании исследований, проведенных Кодреа и Дикой в 2005 году, они определили возраст этих образований как маастрихтско - миоценовый (также предположительно как эггенбургско-оттнангский возраст). Некоторые из редких ископаемых, найденных здесь, также являются позвоночными, и одно из них - хвостовой позвонок завропода . Палеонтологи, участвующие в исследованиях в Рапа Роше, также высказали мнение, что это единственный род завропод, зарегистрированный когда-либо в позднемеловых маастрихтских образованиях в Румынии, который можно было бы назвать Magyarosaurus . [1]
Описание
{kind=link}
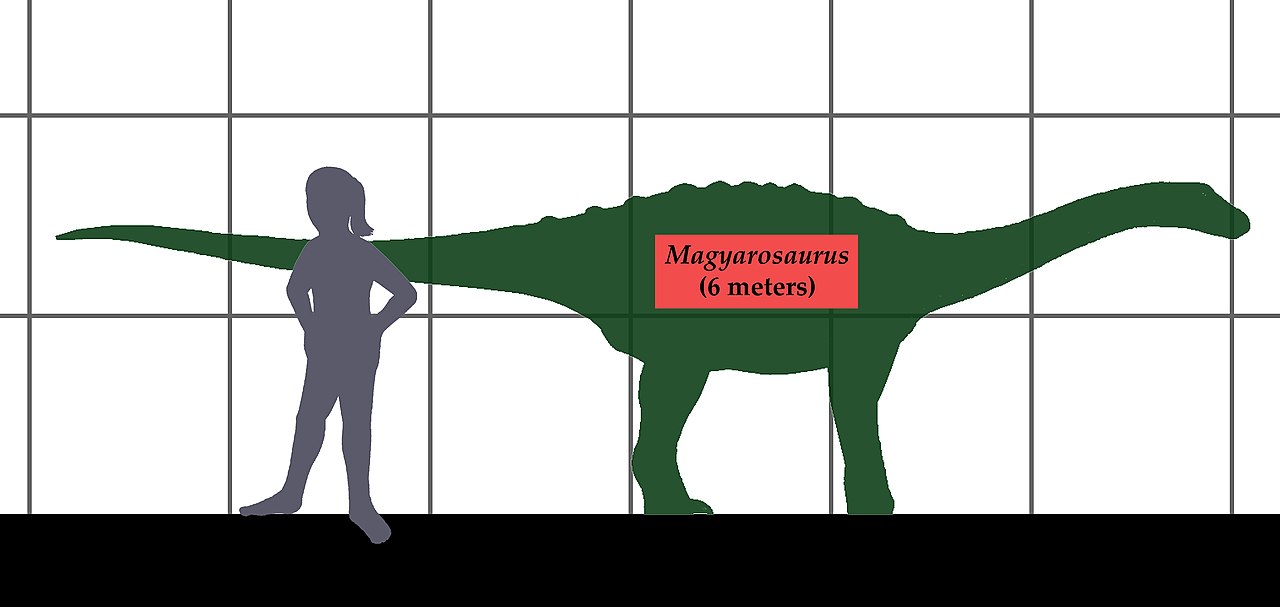
Magyarosaurus был относительно небольшим завроподом, достигавшим 6 м (20 футов) в длину и 750–1000 кг (1650–2200 фунтов) массы тела. [7] [8] Его небольшой размер является отличительной аутапоморфией, поскольку ни один из его близких родственников не имел столь уменьшенного размера. [5] Magyarosaurus также примечателен наличием кожной брони. [9] [10]
Дистальный хвостовой позвонок был отнесен к роду Кодреа и др. (2008). Вероятно, он находился около середины хвоста, поскольку имеет переходные черты. Перед тем, как он был окончательно захоронен, невральная дуга была сломана, вероятно, из-за перемещения позвонков из исходного положения. Его центр удлинен и имеет длину 105 миллиметров (4,1 дюйма). Обе стороны, которые могли бы сочленяться с позвонками, были серьезно повреждены. Он отнесен к Magyarosaurus на основании того, что другие завроподы из региона, в котором он был найден, не известны, и того факта, что он расположен между двумя позвонками по сравнению с ним из-за его промежуточной морфологии. [1]
Палеобиология
Карликовость
Острова, на которых он обитал, привели к тому, что Magyarosaurus стал продуктом островной карликовости в результате селективного давления, представленного ограниченными запасами пищи и отсутствием хищников, что благоприятствовало меньшему размеру тела. [5] Это наблюдается у многих других динозавров, существовавших в то время, включая орнитопода Rhabdodon и нодозавра Struthiosaurus . Nopcsa был первым, кто предложил островную карликовость в качестве объяснения небольшого размера Magyarosaurus по сравнению с другими завроподами. Более поздние исследователи усомнились в его выводах, предположив вместо этого, что известные ископаемые Magyarosaurus представляли собой молодых особей. Однако подробное исследование моделей роста костей, опубликованное в 2010 году, подтвердило первоначальную гипотезу Nopcsa, показав, что маленькие особи Magyarosaurus были взрослыми. [5] [11] Было высказано предположение, что островная карликовость привела к тому, что изолированные роды сохранили более примитивные характеристики. [9]
Гистология
.jpg){kind=link}
.jpg/1280px-Sauropoda_(8030762510).jpg)
В 2010 году Коэн Штейн и др. изучали гистологию Magyarosaurus . Они обнаружили, что даже самые маленькие особи, по-видимому, были взрослыми. Они также сохранили «M. hungaricus » для обозначения более крупных особей, которые были слишком большими, чтобы быть вариациями более мелких особей. Гистология Magyarosaurus показала, что у него была очень низкая скорость роста, но даже при этом он имел высокую скорость метаболизма. [5]
Броня
Остеодерма, обнаруженная в местности "La Cãrare". Местность находится недалеко от деревни Сынпетру, в бассейне Хацег в Румынии . Остеодерма была отнесена к Magyarosaurus dacus . Это показывает, что дермальная броня была широко распространена у этих позднемеловых завропод. [10] Остеодерма имела своеобразную форму и размер, [10] что привело к тому, что яйца были отнесены к его семейству Nemegtosauridae и, возможно, к Magyarosaurus . [9]
Возможные яйца
{kind=link}

Яйца литостротий были отнесены к Nemegtosauridae . Яйца, возможно, принадлежат либо Magyarosaurus dacus , либо Paludititan , причем первый вариант более вероятен. [9] Бассейн Хацег был крупным местом гнездования в позднем меловом периоде и служил местом обитания гадрозавридов и титанозавров. 11 яиц были отнесены к Nemegtosauridae , все из формации Санпетру. [9] Внутри яиц сохранились эмбрионы, и одно яйцо демонстрирует доказательства наличия кожной брони. [9]
Яйца были обнаружены в 2001 году во время полевой экспедиции бельгийско-румынской команды. Первоначально они были идентифицированы как находящиеся в гнездах, но теперь было показано, что никаких гнездовых структур не сохранилось. [9]
Палеоэкология
В раннем маастрихте бассейн Хацег был субгумидным и имел сезонные осадки. Однако в более поздний период формирования произошло крупномасштабное изменение палеоэкологии, регион превратился в обширную водно-болотную угодье. [12]
Magyarosaurus dacus известен из раннего маастрихта формации Санпетру, части бассейна Хацег в Румынии. [12] [2] [13] Из бассейна Хацег также известны небольшие базальные гадрозавриды Telmatosaurus ; [5] небольшой нодозаврид Struthiosaurus ; [14 ] манирапторы Balaur , Bradycneme и Elopteryx ; [14 ] птерозавр Хацегоптерикс ; [11] [15] и два вида эуорнитоподов Zalmoxes . [5]
M. sp. известен из позвонка. Позвонок был найден в позднем мелу формации Себеш , хотя он, вероятно, был вымыт из формации Шард и помещен туда. Наряду с Magyarosaurus существовал Kallokibotion , древняя черепаха ; [1] Balaur , двукоготный авиалан ; [14] и Eurazhdarcho , аждархид . [15] Наряду с Magyarosaurus , Telmatosaurus и Zalmoxes также являются карликовыми родами, что доказано их гистологией. [5]
Смотрите также
Ссылки
- ^ abcd Кодря, Вирджиния; Мурзеа-Джипа, К.; Венцель, М. (2008). «Позвонки зауроподов в Рапа Рошье (район Альба)» (PDF) . Acta Palaeontologica Romane . 6 : 43–48 . Архивировано из оригинала (PDF) 3 марта 2016 г. Проверено 28 января 2014 г.
- ^ ab Карри Роджерс, К. (2005). "Титанозавры: филогенетический обзор" в Карри Роджерс, К. и Уилсон, JA (редакторы), Зауроподы: эволюция и палеобиология . Беркли: Издательство Калифорнийского университета. ISBN 0-520-24623-3
- ^ Нопца, Ф (1915). «Die Dinosaurier der siebenburgischen Landesteile Ungarns». Унгар. геол. Рейхсанст . 23 : 1–26 .
- ^ фон Хюэн, Ф. (1932). «Die ископаемые Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte». Монг. геол. Пал. , 4 (1) балла. 1 и 2, viii +361 стр.
- ^ abcdefgh Stein, K.; Csiki, Z.; Curry Rogers, K.; Weishampel, DB; Redelstorff, R.; Carballidoa, JL; Sander, PM (2010). «Маленький размер тела и экстремальное ремоделирование кортикальной кости указывают на филетическую карликовость у Magyarosaurus dacus (Sauropoda: Titanosauria)». Труды Национальной академии наук Соединенных Штатов Америки . 20. 107 (20): 9258– 9263. Bibcode : 2010PNAS..107.9258S. doi : 10.1073/pnas.1000781107 . PMC 2889090. PMID 20435913 .
- ^ «Брифинг», Geology Today 7 (1): стр. 2-6.
- ^ Пол, ГС (2010) Принстонский полевой путеводитель по динозаврам , Princeton University Press, стр. 213
- ^ Бенсон, Роджер Б. Дж.; Кампионе, Николас Э.; Каррано, Мэтью Т.; Мэннион, Филип Д.; Салливан, Корвин; Апчерч, Пол; Эванс, Дэвид К. (2014-05-06). «Темпы эволюции массы тела динозавров указывают на 170 миллионов лет устойчивых экологических инноваций в родословной птиц». PLOS Biology . 12 (5): e1001853. doi : 10.1371/journal.pbio.1001853 . ISSN 1545-7885. PMC 4011683. PMID 24802911 .
- ^ abcdefg Греллет-Тиннер, Г; Кодреа, В; Фоли, А; Хига, А.; Смит, Т. (2012). Эндрю А. Фарке (ред.). «Первые доказательства репродуктивной адаптации к «островному эффекту» карликового мелового румынского титанозавра с эмбриональным покровом в яйце». PLOS ONE . 7 (3): e32051. Bibcode :2012PLoSO...732051G. doi : 10.1371/journal.pone.0032051 . PMC 3297589 . PMID 22412852.
- ^ abc Csiki, З. (1999). «Новые свидетельства существования панцирных титанозавридов в позднем мелу - Magyarosaurus dacus из бассейна Хатег (Румыния)». Ориктос . 2 : 93–99 .
- ^ ab Скотт, К. (2012). «Изменение кубика» . В МакАртур, К.; Рейал, М. (ред.). Планета динозавров . Firefly Books. стр. 200–208. ISBN 978-1-77085-049-1.
- ^ ab Therrien, F.; Zelenitsky, DK; Weishampel, DB (2009). «Палеоэкологическая реконструкция позднемеловой формации Санпетру (бассейн Хацег, Румыния) с использованием палеопочв и ее значение для «исчезновения» динозавров». Палеогеография, Палеоклиматология, Палеоэкология . 272 ( 1–2 ): 37–52 . Bibcode : 2009PPP...272...37T. doi : 10.1016/j.palaeo.2008.10.023.
- ^ B. Vila; A. Galobart; JU Canudo; J. Le Loeff; et al. (2012). «Разнообразие динозавров-завропод и их первая таксономическая последовательность из позднего мела юго-западной Европы: ключи к гибели и вымиранию». Палеогеография, Палеоклиматология, Палеоэкология . 350–352 (15): 19–38 . Bibcode : 2012PPP...350...19V. doi : 10.1016/j.palaeo.2012.06.008.
- ^ abc Weishampel, DB; Jianu, CM (2011). Трансильванские динозавры . Издательство Университета Джона Хопкинса. С. 37–38 . ISBN 978-1-4214-0027-3.
- ^ ab Vremir, MTS; Kellner, AWA; Naish, D.; Dyke, GJ (2013). Viriot, Laurent (ред.). "Новый птерозавр Azhdarchid из позднего мела Трансильванского бассейна, Румыния: последствия для разнообразия и распространения Azhdarchid". PLOS ONE . 8 (1): e54268. Bibcode :2013PLoSO...854268V. doi : 10.1371/journal.pone.0054268 . PMC 3559652 . PMID 23382886.
