Лисовица
| Лисовица Временной диапазон: поздний триас | |
|---|---|
 | |
| Левая плечевая и бедренная кость | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Анималия |
| Тип: | Хордовые |
| Клад : | Синапсида |
| Клад : | Терапсида |
| Подотряд: | † Аномодонтия |
| Клад : | † Дицинодонтии |
| Семья: | † Stahleckeriidae |
| Подсемейство: | † Плацериевые |
| Род: | † Лисовица Сулей и Недзведски, 2019 г. |
| Разновидность: | † Л. бояни |
| Биномиальное имя | |
| † Лисовица бояни Сулей и Недзведски, 2019 | |
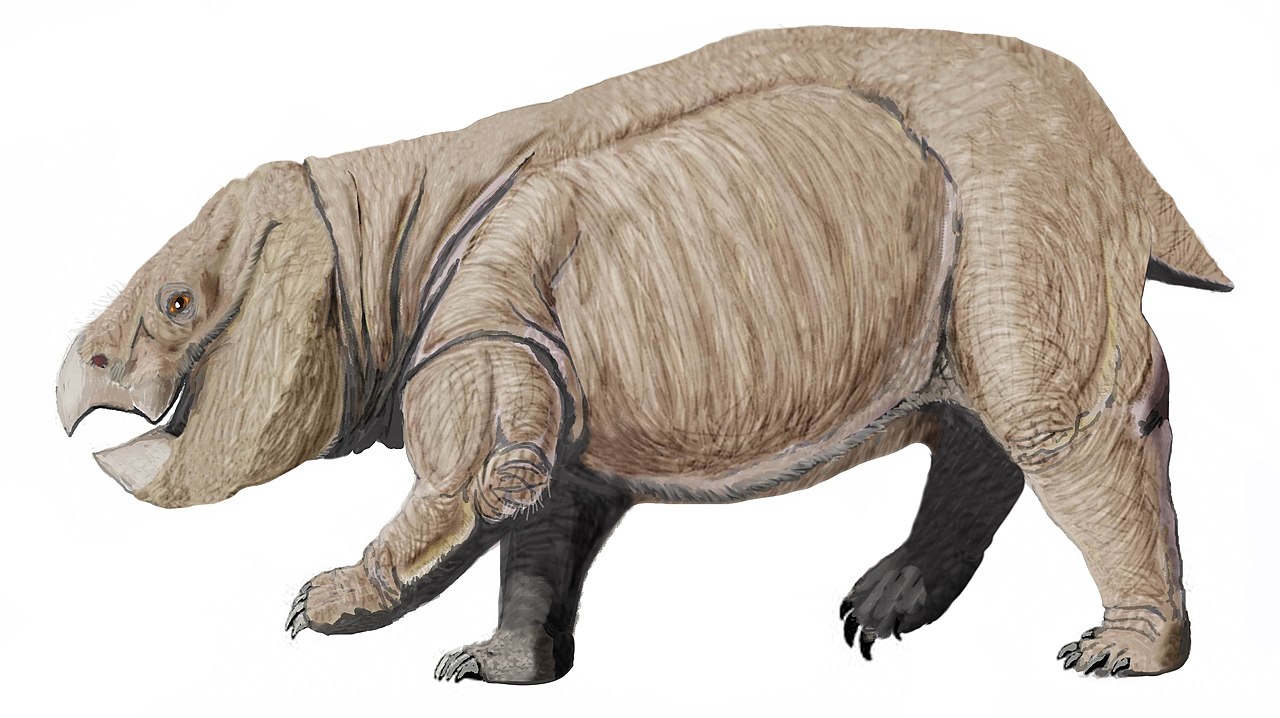
Lisowicia — вымерший род гигантских дицинодонтов- синапсидов , которые обитали на территории современной Польши в поздненорийскомили раннеретийском ярусах позднего триасового периода , около 210–205 миллионов лет назад. Lisowicia — крупнейший известный дицинодонт, а также крупнейший не млекопитающий синапсид, достигающий около 4,5 метров (15 футов) в длину, до 2,6 метров (8,5 футов) в высоту в бедрах и весящий около 5–7 метрических тонн (5,5–7,7 коротких тонн), что сопоставимо по размеру с современными слонами . Он также был одним из последних дицинодонтов, живших незадолго до своего вымирания в конце триасового периода. Ископаемые останки гигантского дицинодонта были известны из Польши с 2008 года, но Lisowicia не была названа и официально описана как новый вид до конца 2018 года.
Lisowicia уникальна среди дицинодонтов своей прямой осанкой, когда все четыре конечности находятся прямо под ее телом. Это похоже на конечности современных млекопитающих и динозавров , но в отличие от раскинувшихся и полупрямых поз, типичных для всех других дицинодонтов (и, конечно, всех других не млекопитающих синапсид), и разделяет многие независимо эволюционировавшие черты ее конечностей с крупными млекопитающими. Lisowicia была самым крупным животным в своей среде обитания и была сопоставима по размеру и экологии с травоядными «прозавроподами» зауроподоморфами , которые в остальном занимали роль крупных высокорослых травоядных в большинстве экосистем позднего триаса. До открытия Lisowicia предполагалось, что дицинодонты были вытеснены зауроподоморфами и не могли достичь столь же больших размеров тела. Lisowicia могла развить свой большой размер в ответ на отсутствие завроподоморфов в своей экосистеме на юге Польши или же могла выступать в качестве их прямого конкурента.
Описание

Хотя Lisowicia была очень крупной для дицинодонта, она все еще очень похожа на других триасовых stahleckeriids , таких как ее близкий родственник Placerias из Северной Америки. Это было крепко сложенное четвероногое животное с округлым, бочкообразным телом, большой головой с клювовидными челюстями и короткой шеей, коренастыми конечностями и очень коротким хвостом. [1]
Череп неполный и известен только по разным изолированным отдельным частям, но их достаточно, чтобы определить, что у него был череп, похожий на череп других stahleckeriids. Височные окна были большими, а задняя часть черепа была втянута в высокий гребень между ними, обеспечивая обширное прикрепление челюстных мышц в задней части черепа. Морда была относительно удлиненной, как у других stahleckeriids, и имела беззубый черепахоподобный клюв спереди. Как и некоторые другие триасовые дицинодонты, Lisowicia была полностью беззубой и не имела даже бивней, типичных для большинства дицинодонтов. Вместо этого у нее была пара коротких и толстых треугольных выступов от верхней челюстной кости позади клюва, называемых клыкообразными отростками, похожими на таковые у родственной Ischigualastia и других stahleckeriids. Аналогичным образом, сравнение пропорций квадратной кости в задней части черепа с пропорциями Ischigualastia позволяет предположить, что у Lisowicia был широкий череп, ширина которого между каждой квадратной костью в челюстных суставах составляла примерно 43 сантиметра (17 дюймов). [2]
Его позвоночник и ребра типичны для дицинодонтов, однако шейные позвонки шеи примечательны необычными углублениями, вырытыми по бокам. Это похоже на отверстия в некоторых позвонках динозавров, которые связаны с системой воздушных мешков и дыхательной системой, подобной птичьей . Однако, в отличие от динозавров, у Lisowicia отсутствуют отверстия в позвонках ( плевроцели ) для воздушных мешков, а внутренняя часть позвонка сплошная. Эта черта уникальна для Lisowicia среди дицинодонтов, и ее функция неясна. Кости таза и задних конечностей очень прочные по сравнению с другими дицинодонтами из-за огромного размера животного (например, самая большая известная бедренная кость имеет длину 80 сантиметров (31 дюйм)), но в остальном похожи на кости других stahleckeriids. Как и у других stahleckeriids, задние ноги также удерживались вертикально под телом. [1]
Передние конечности
Передние конечности Lisowicia очень характерны для дицинодонтов. Действительно, их конструкция уникальна не только для группы, но и для не млекопитающих синапсидов в целом и разделяет черты с конечностями млекопитающих и динозавров. Передние конечности Lisowicia удерживаются полностью прямо под телом, с локтевым суставом, который направлен назад и поэтому позволяет предплечью качаться вперед и назад только при парасагиттальной походке. Такое расположение встречается в задних конечностях различных других крупных триасовых kannemeyeriiforms , но все они сохраняют раскинутые передние конечности с локтями, согнутыми наружу, и суставами, которые позволяют лучевой и локтевой костям вращаться во время каждого шага в раскинутой позе.
Чтобы достичь этой позы, передние конечности Lisowicia должны были претерпеть несколько заметных анатомических изменений по сравнению с нормальным состоянием у дицинодонтов. На плечевой кости плечевой сустав и локоть параллельны друг другу, в отличие от повернутой плечевой кости раскинувшихся дицинодонтов, и поэтому плечевая кость удерживается вертикально под плечом и прямо над лучевой и локтевой костями . Само предплечье также необычно короткое по сравнению с другими дицинодонтами, поскольку локоть теперь расположен ниже прямо под телом и ближе к земле. Мускулатура передней конечности также была перестроена, чтобы облегчить ее вертикальную позу и походку, теперь она функционирует для того, чтобы тянуть конечность вперед и назад и теряет способность вращать верхнюю часть руки, как это было бы при раскинутом шаге. [1]
Размер

Хотя скелет неполный, массивный размер различных элементов его скелета указывает на то, что Lisowicia была очень крупным животным, крупнее любого другого дицинодонта. Сравнивая его кости с костями других дицинодонтов, длина его тела оценивается примерно в 4,5 метра (15 футов), а высота бедра — в 2,6 метра (8,5 фута). [3] [4] [1] Эта верхняя оценка соответствует самой большой сохранившейся частичной плечевой кости, которая в полном виде оценивается в 61,5 сантиметра (24,2 дюйма) в длину на основе пропорций полных плечевых костей. [5] Первоначально вес его тела был рассчитан примерно на 9,3 метрических тонны (10,3 коротких тонны) из окружности его бедренной и плечевой костей — приблизительный показатель для оценки того, какой вес должны были поддерживать конечности. Однако стержни этих костей у Lisowicia непропорционально крепкие и толстые , даже для его размера, поэтому его масса тела, вероятно, была завышена. [1] [6]
Его вес тела был позже более надежно оценен путем моделирования его массы из предполагаемого общего объема его тела (объемное моделирование). Эти оценки варьировались в зависимости от обхвата его грудной клетки и количества мягких тканей, смоделированных вокруг скелета, с общим средним весом 5,9 метрических тонн (6,5 коротких тонн), и самой низкой оценкой с минимальным содержанием жира и других тканей в теле 4,9 метрических тонн (5,4 коротких тонн) и максимальной 7 метрических тонн (7,7 коротких тонн) в самом крупном его размере. Этот диапазон сопоставим с диапазоном современных африканских слонов , что делает Lisowicia крупнейшим известным немлекопитающим синапсидом и самым большим синапсидом до появления крупных млекопитающих в эоцене (таких как бронтотерии ). Для сравнения, крупный диноцефальный Tapinocaninus был оценен в среднем всего в 892 килограмма (1967 фунтов) с использованием тех же методов. [6] [7]
История открытия

Останки Lisowicia были обнаружены в глиняном карьере Липе-Слёнске в Лисовице , деревне на юге Польши недалеко от города Люблинец , Силезия . Ископаемые слои Липе-Слёнске состоят в основном из зеленых аргиллитов . Возраст глиняного карьера Липе-Слёнске было трудно определить, поскольку различные биостратиграфические методы датирования давали разные результаты. Обычно местонахождение датировалось ретским ярусом , что сделало бы Lisowicia одним из самых молодых дицинодонтов в мире. Однако палеонтологи Гжегож Рацкий и Спенсер Лукас заявили, что местонахождение старше и датируется средним норийским ярусом на основе биостратиграфии крупных позвоночных, включая Lisowicia . Вместо этого они предположили, что Lisowicia была не намного моложе других дицинодонтов и была близка по возрасту к последним известным родам в других местах в мире. [8]
Образец детритовых цирконов из глиняного карьера Липе-Слёнске был радиометрически датирован для определения максимального возраста отложений ископаемых пластов. Самое молодое зерно циркона имело возраст приблизительно 211 ± 3 миллиона лет в позднем норийском яме, однако из-за несоответствия между образованием кристаллов циркона и отложением в виде осадка, вероятно, что Липе-Слёнске моложе этого возраста, что подтверждает оценки от позднего норийского яруса до раннего ретийского яруса. [9]
_Lisowicia_bojani.jpg/1280px-Szkic_wst%C4%99pnej_rekonstrukcji_(2008)_Lisowicia_bojani.jpg)
Останки Lisowicia были впервые обнаружены в 2006 году палеонтологами Ежи Дзиком, Томашем Сулеем и Гжегожем Недзведским, которые изначально приняли их за кости динозавра-зауроподоморфа из-за их огромных размеров. Позже, в 2008 году, они распознали окаменелости как останки гигантского дицинодонта, к тому времени известного по частично сочлененному скелету и различным изолированным костям по крайней мере двух особей. В период с 2007 по 2014 год из глиняной ямы было собрано более 1000 костей Lisowicia . [10] [2] [11]
Lisowicia не будет официально названа и опубликована как новый таксон до января 2019 года, хотя название и описание были опубликованы в сети в ноябре 2018 года до официальной даты публикации. Род назван в честь деревни Лисовице, где были найдены оригинальные образцы, в то время как видовое название bojani дано в честь Людвига Генриха Боянуса (1776–1827), немецкого врача и натуралиста , который провел пионерскую работу в области сравнительной анатомии и палеонтологии . [10] Известные ископаемые останки Lisowicia хранятся в коллекциях Института палеобиологии Польской академии наук и кафедры палеобиологии и эволюции Варшавского университета , Польша.
Классификация
Lisowicia — это kannemeyeriiform, группа, к которой принадлежит большинство триасовых дицинодонтов, и является членом семейства Stahleckeriidae , единственной оставшейся линии дицинодонтов, присутствующей в позднем триасе. В частности, филогенетический анализ, проведенный Sulej & Niedźwiedzki (2019) с использованием двух отдельных наборов данных, показал, что Lisowicia тесно связана с известным североамериканским родом Placerias в подсемействе Placeriinae , вместе с марокканским Moghreberia . Аналогичный результат был получен Szczygielski & Sulej (2023) при описании польского дицинодонта Woznikella , которые также обнаружили южноафриканского Pentasaurus в основании Placeriinae, в то время как сам Woznikella был найден в более отдаленном положении за пределами Stahleckeriidae. [12]
Приведенная ниже кладограмма представляет собой измененную и упрощенную версию работы Щигельского и Сулей (2023), которая фокусируется на взаимоотношениях стахлеккериид и их ближайших родственников и отображает строгий консенсус трех самых коротких ( самых экономных ) деревьев: [12]




Альтернативная некладистическая филогения для kannemeyeriiforms была предложена Томашем Сулеем в 2024 году в его остеологическом описании Lisowicia . Вместо того, чтобы определять отношения, основанные только на морфологии, как в большинстве кладистических анализов, Сулей также использовал стратиграфическую и биогеографическую информацию для выдвижения гипотез о взаимоотношениях предков и потомков в пространстве и времени. Этот подход в палеонтологии был назван «хронофилетикой» польским палеонтологом Ежи Дзиком . [13] Эти результаты заметно отличаются от традиционных филогений, основанных на кладистике, таких как дерево, показанное выше. Сулей определил Lisowicia как последнего члена лавразийской линии kannemeyeriiform, начинающейся с «kannemeyeriid» Shaanbeikannemeyeria в среднем триасе , через Rabidosaurus , Woznikella (в отличие от его более отдаленного положения, восстановленного кладистически) к Placerias, прежде чем достичь кульминации в Lisowicia . В отличие от кладистического дерева выше, он предположил, что большинство stahleckeriines не имели близкого общего предка с placeriines, а вместо этого произошли от отдельной линии среднетриасовых «kannemeyeriid», которые мигрировали в Гондвану, спускаясь от лавразийских Rhadiodromus и Parakannemeyeria . [5]
Сулей утверждал, что форма лобных и теменных костей, особенно их поперечное сечение вдоль средней линии, где они окружают мозговую коробку, была одним из ключевых отличий, разделяющих эти линии происхождения. У Lisowicia и членов лавразийской линии теменные кости вертикально удлиненные и треугольные, сужающиеся книзу к точке. Напротив, у stahleckeriines и членов их линии теменные кости короче, но имеют более широкие, более округлые нижние края. На поверхности теменное отверстие также расположено ближе к задней части черепа в лавразийской линии, а лобные кости длиннее и участвуют в крае глазницы. Единая сочленовная поверхность для плеча и ребер на грудине также была идентифицирована как характеристика этой линии. [5]
Дицинодонтный гигантизм и эволюция

До открытия Lisowicia считалось, что эволюция гигантских размеров тела у триасовых завроподоморфов является исключительной чертой динозавров, поскольку другие крупные травоядные, включая дицинодонтов, не считались достигшими подобных размеров. Слоновий размер Lisowicia показывает, что гигантизм также развился у дицинодонтов, а анализ размера их тела по мере их эволюции в течение позднего триаса показывает, что, как и завроподоморфы, триасовые дицинодонты также увеличивались в размерах тела, достигнув кульминации в Lisowicia . Фактически, дицинодонты были последовательно крупнее завроподоморфов в течение карнийского яруса, и оба достигли схожих гигантских размеров в норийском и ретийском ярусах с Lisowicia . Эволюция гигантизма как у зауроподоморфов, так и у Lisowicia в схожие периоды времени предполагает, что экологические факторы могли позволить и побудить обе линии вырасти до гигантских размеров, в отличие от специфических черт, присущих динозаврам-зауроподоморфам, как предполагалось ранее. [1]
Эволюция гигантизма у дицинодонтов была постепенным процессом, который длился около 20 миллионов лет и приобрел серьезную перестройку опорно-двигательного аппарата для достижения размера и осанки Lisowicia . Возможно, что гигантизм у Lisowicia был вызван увеличением размера тела у хищников или метаболической адаптацией для получения большего количества питательных веществ из пищи за счет увеличения размера кишечника. [14] [15] Это также подчеркивает, что эволюция дицинодонтов в позднем триасе все еще плохо изучена. Ранее считавшаяся географически ограниченной и находящейся в упадке, Lisowicia демонстрирует, что дицинодонты все еще были географически широко распространены и продолжали диверсифицироваться в позднем триасе, в частности, развивая черты, поддерживающие огромные размеры тела. [1]
Палеобиология
Кормление и диета
Lisowicia была травоядным животным, как и большинство других дицинодонтов, использовавшим свой относительно длинный клюв для сбора и жевания, хотя его большой размер подразумевает, что он питался более высокими уровнями растительности, чем это было типично для группы. С другой стороны, Сулей (2024) предположил, что Lisowicia была «травоядным», питавшимся низкорасположенной растительностью. Это предположение основывалось на ориентации глазниц и низком положении затылочных мыщелков (где череп соединяется с шеей), что интерпретировалось как указание на то, что его голова обычно держалась наклоненной вниз к земле. В этом положении слегка направленные вверх глазницы теперь были бы обращены глазами вперед к горизонту во время еды. [5]
Копролиты, приписываемые Lisowicia, указывают на то, что он в основном питался мягкой растительностью и хвойными . Однако некоторые копролиты содержат большие порции древесного материала, черта, наблюдаемая у некоторых других копролитов дицинодонтов. Было высказано предположение, что это отражает сезонные изменения в его рационе, возможно, когда их предпочитаемая диета из мягкой растительности была вне сезона. [14]
Хищничество

Многочисленные следы укусов были обнаружены на костях молодых дицинодонтов из той же местности, где были найдены дицинодонт Lisowicia и хищный архозавр Smok , что указывает на то, что молодые дицинодонты были предпочтительно и активно объектом охоты Smok . Фрагменты костей дицинодонтов также были обнаружены в предполагаемых копролитах Smok , что подразумевает, что дицинодонты, такие как Lisowicia, были основным компонентом в рационе хищника. [15] [16] Основываясь на отсутствии носовых костей и редкости других костей морды, Сулей (2024) предположил, что морда Lisowicia могла быть структурно слабой и поэтому предпочтительно подвергалась нападению хищников, таких как Smok, либо во время хищничества, либо путем отчленения морды от туши. [5]
Рост
Гистологические исследования костей конечностей Lisowicia показали черты, характерные для быстрого роста молодых особей, как и у других крупных дицинодонтов. Однако, в то время как скорость роста других дицинодонтов замедлялась по мере их роста, изученные кости Lisowicia не показали таких признаков замедления роста. Возможность того, что кости принадлежали молодым особям с еще большим размером тела взрослой особи, чем предполагалось, была исключена частично на основании их уже огромного размера, но также и потому, что кости были хорошо окостеневшими и зрелыми в плане развития. Вместо этого более вероятно, что Lisowicia росла очень быстро без каких-либо перерывов, пока не достигла своего взрослого размера тела, поскольку не было никаких четких линий остановки роста (LAG), которые в противном случае указывали бы на периоды более медленного, сниженного роста. LAG могли действительно отсутствовать или, возможно, были стерты обширной ремоделировкой кости во взрослом возрасте, и то и другое поддерживало бы постоянный быстрый рост Lisowicia , подобно млекопитающим и некоторым динозаврам. [1] [10]
Туалеты
Копролиты, приписываемые Lisowicia, были обнаружены в обильных скоплениях в областях, которые, как полагают, были вокруг стоячей воды. Эти скопления похожи на находки в Бразилии, которые, как полагают, представляют собой общественные туалеты, сделанные дицинодонтом Dinodontosaurus , и вполне возможно, что Lisowicia могла вести себя похожим образом. Общественные туалеты задокументированы у современных стадных млекопитающих и подтверждают идею о том, что дицинодонты, такие как Lisowicia, жили стадами, и даже предполагают, что у них было сложное социальное поведение, как у современных крупных млекопитающих. [14] [17]
Палеоэкология

Lisowicia сосуществовала с широким спектром позднетриасовых животных в Липе-Слёнске. Большинство этих животных были архозаврами, включая крупного хищного архозаврида Smok , небольшими динозавроморфами, включая силезаврида и целофизиподобного неотеропода , птерозаврами и небольшим крокодиломорфом (ранее считавшимся попозавроидом ) . Другие мелкие рептилии включают сфенодонта , неопределенного архозавроморфа и возможного хористодера . Единственным другим синапсидом, который , как известно, сосуществовал с Lisowicia, был Hallautherium , небольшой морганукодонтный млекопитающий, тесно связанный с современными млекопитающими и на много порядков меньший, чем Lisowicia . Влажная, пресноводная среда обитания поддерживала темноспондильные амфибии ( капитозавр и более мелкие плагиозавры ), а также обилие рыб, включая двоякодышащих рыб , латимерий и акул -гибодонтов . [8] [16]
Окружающая среда в Липе-Слёнске была влажной и болотистой, сравнимой с современными болотами , [14] с медленно текущими извилистыми реками и старицами , окруженными обильной растительностью, включающей хвойные деревья семейства Cheirolepidiaceae и гинкговые , семенные папоротники , такие как Lepidopteris и саговники Androstrobus и Beania . Напротив, соседние среды могли быть сухими и засушливыми, что подтверждается остатками, возможно, ксерофитного хвойного дерева Hirmeriella и фрагментами древесного угля от пожаров, которые переносились реками в эту местность. [2] [18] [19]
Отсутствие крупных травоядных зауроподоморфов любопытно, поскольку они присутствуют во многих других экосистемах позднего триаса, в том числе в близлежащих местах в Европе, например, Plateosaurus в Германии. Возможно, что гигантские дицинодонты, такие как Lisowicia, напрямую конкурировали с зауроподоморфами, однако также возможно, что их отсутствие не было связано, и вместо этого Lisowicia выросла до своих огромных размеров, чтобы заполнить экологическую роль крупных травоядных, которые в других экосистемах занимали зауроподоморфы. [3]
Ссылки
- ^ abcdefgh Sulej, Tomasz; Niedźwiedzki, Grzegorz (4 января 2019 г.). «Позднетриасовый синапсид размером со слона с прямыми конечностями». Science . 363 (6422): 78– 80. Bibcode :2019Sci...363...78S. doi : 10.1126/science.aal4853 . ISSN 0036-8075. PMID 30467179.
- ^ abc Дзик, Ежи; Сулей, Томаш; Недзведский, Гжегож (2008). «Ассоциация дицинодонтов-тероподов в последнем триасе Польши». Acta Palaeontologica Polonica . 53 (4): 733–738 . doi : 10.4202/app.2008.0415 .
- ^ ab Дворски, Джордж (22 ноября 2018 г.). «Во время триаса этот родственник млекопитающих размером со слона жил рядом с динозаврами». Gizmodo . Получено 10 сентября 2022 г. .
- ↑ Сотрудники (23 ноября 2018 г.). «Обнаружен двоюродный брат гигантского млекопитающего». Science Daily . Архивировано из оригинала 22 августа 2022 г. Получено 10 сентября 2022 г.
- ^ abcde Сулей, Т. (2024). «Остеология и взаимоотношения позднетриасового гигантского дицинодонта Lisowicia ». Зоологический журнал Линнеевского общества . 202 (1). zlae085. doi : 10.1093/zoolinnean/zlae085.
- ^ ab Romano, Marco; Manucci, Fabio (14 июня 2019 г.). «Изменение размеров Lisowicia bojani : оценка объемной массы тела и трехмерная реконструкция гигантского позднетриасового дицинодонта». Historical Biology . 33 (4): 474– 479. doi : 10.1080/08912963.2019.1631819. ISSN 0891-2963. S2CID 196679837.
- ^ Романо, Марко; Рубидж, Брюс (2019). «Первая 3D-реконструкция и оценка объемной массы тела тапиноцефалидного диноцефала Tapinocaninus pamelae (Synapsida: Therapsida)». Историческая биология . 33 (4): 498– 505. doi : 10.1080/08912963.2019.1640219. ISSN 1029-2381. S2CID 203881268.
- ^ ab Racki, G.; Lucas, SG (2018). «Время вымирания дицинодонтов в свете необычной польской фауны позднего триаса и подход Кювье к вымиранию». Историческая биология . 32 (4): 1– 11. doi :10.1080/08912963.2018.1499734. S2CID 91926999.
- ^ Kowal-Linka, M.; Krzemińska, E.; Czupyt, Z. (2019). «Самые молодые детритовые цирконы из континентальных отложений верхнего триаса Lipie Śląskie (Lisowice) (Польша): Последствия для максимального возраста осадконакопления костеносного горизонта Lisowice». Палеогеография, Палеоклиматология, Палеоэкология . 514 : 487– 501. Bibcode : 2019PPP...514..487K. doi : 10.1016/j.palaeo.2018.11.012. S2CID 134592379.
- ^ abc Гретхен Фогель (22 ноября 2018 г.). «Гигант во времена динозавров: древний двоюродный брат млекопитающего выглядел как нечто среднее между носорогом и черепахой». Science . doi :10.1126/science.aaw1183. S2CID 135360687 . Получено 22 декабря 2019 г. .
- ^ Сент-Флер, Николас (4 января 2019 г.). «Родственник млекопитающих размером со слона, пасшийся рядом с динозаврами». The New York Times . Получено 6 января 2019 г.
- ^ аб Щигельски, Т.; Сулей, Т. (2023). «Woznikella triradiata n. gen., n. sp. - новый каннемейериформный дицинодонт из позднего триаса северной Пангеи и глобальное распространение триасовых дицинодонтов». Comptes Рендус Палевол . 22 (16): 279–406 . doi : 10.5852/cr-palevol2023v22a16 .
- ^ Дзик, Дж. (2005). «Хронофилетический подход: стратофенетика перед лицом неполной палеонтологической летописи» (PDF) . Специальные статьи по палеонтологии . 73 : 159–183 .
- ^ abcd Bajdek, P.; Owocki, K.; Niedźwiedzk, G (2014). «Предполагаемые копролиты дицинодонтов из верхнего триаса Польши». Палеогеография, Палеоклиматология, Палеоэкология . 411 : 1– 17. Bibcode :2014PPP...411....1B. doi :10.1016/j.palaeo.2014.06.013.
- ^ ab Niedźwiedzki, G.; Gorzelak, P.; Sulej, T. (2011). «Следы укусов на костях дицинодонтов и ранняя эволюция крупных наземных хищников». Lethaia . 44 (1): 87– 92. Bibcode : 2011Letha..44...87N. doi : 10.1111/j.1502-3931.2010.00227.x.
- ^ ab Qvarnström, Martin; Ahlberg, Per E.; Niedźwiedzki, Grzegorz (2019). «Остеофагия триасового архозаврида, подобная тираннозавридовой». Scientific Reports . 9 (1): 925. Bibcode :2019NatSR...9..925Q. doi :10.1038/s41598-018-37540-4. PMC 6353991 . PMID 30700743.
- ^ Фиорелли, Ле; Эскурра, доктор медицины; Хехенляйтнер, ЕМ; Арганьярас, Э; Таборда, JRA; Троттейн, MJ; фон Бачко, MB; Дезохо, JB (2013). «Самые старые известные общественные уборные свидетельствуют о стадизме у мегатравоядных триасовых животных». Научные отчеты . 3 (3348): 3348. Бибкод : 2013NatSR...3.3348F. дои : 10.1038/srep03348 . ПМЦ 3842779 . ПМИД 24287957.
- ^ Jewuła, K.; Matysik, M.; Paszkowski, M.; Szulc, J. (2019). «Позднетриасовое развитие плайи, поймы гильгай и флювиальных сред Верхней Силезии, южная Польша». Sedimentary Geology . 379 : 25–45 . Bibcode : 2019SedG..379...25J. doi : 10.1016/j.sedgeo.2018.11.005. ISSN 0037-0738. S2CID 133862052.
- ^ Wawrzyniak, Z. (2010). «Чем могли питаться дицинодонты (Dicynodontia, Therapsida) из Липе-Слёнске? (Верхняя Силезия, Польша) – предварительные результаты исследования макрофлоры позднего триаса». В Nowakowski, D. (ред.). Морфология и систематика ископаемых позвоночных . DN Publisher, Вроцлав. С. 118–124 . OCLC 751467953.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
_Lisowicia_bojani.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
